Où l'on retrouve les empreintes digitales ?
Les empreintes digitales 

Évidemment, en insistant sur l’existence de deux modes de croissance, je simplifie à outrance. Pour chacun de ces systèmes, il existe un ensemble de conditions qui vont donner des plis ou des branches plus ou moins marqués, plus ou moins denses, ou pas de plis ni de branches. Il y a, dans le jargon de la morphogenèse, des « diagrammes de morphologie » ; aux marges de ces diagrammes, on quitte complètement les mécanismes que j’ai évoqués, ou bien on les noie dans autre chose.
En outre, les mécanismes en question peuvent être camouflés très astucieusement, et il faut alors de l’imagination ou de la chance pour les débusquer. Ainsi, les empreintes digitales elles-mêmes (nous y revoilà) sont attribuées depuis longtemps (depuis les années 1920, à peu près) à un mécanisme de formation de plis, et tous les chercheurs du domaine (et du domaine connexe de la police scientifique) répètent en chœur le même credo depuis soixante-dix ans. On voit souvent des plis réguliers analogues sur le goudron des boulevards, par exemple sur la promenade des Anglais, à Nice, autour des bouches d’égout. J’en ai vu aussi dans le quartier du Marais, à Paris, près du métro Pont-Marie.
Mais croire que les empreintes digitales se forment par plissement, c’est oublier, ou négliger un article de 1951 de Alfred Haie, un embryologiste qui a essayé de voir si les empreintes digitales étaient liées à des plis. Si ça plie, il doit y avoir un déplacement des couches, de sorte que certaines parties montent et d’autres descendent ; c’est la nature même du pli. Or il ne décrit dans ces travaux rien de tel ; on peut montrer qu’il y a peut-être des forces ayant tendance à plier, mais les empreintes ne se forment pas par pli.
Qu’il y ait des forces pouvant provoquer des plis, cela ne fait guère de doute. C’est à cause des coussinets, dont il a été question au premier chapitre : en se dégonflant, les coussinets tendent à faire des plis, un peu comme un pruneau d’Agen qui sèche. Le problème c’est que… ça ne plie pas. Ce qu’on voit, ce sont des différences de vitesse de croissance, en sorte que ça se développe plus ici, moins là, et au final on dirait que ça a plié, sans plier.
On pourrait tout aussi bien jeter aux orties le mécanisme de pli. Mais on le rattrape in extremis par la culotte, car en fait, il n’est pas nécessaire que la chose plie pour faire des plis. Ce paradoxe apparent trouve sa solution dans ce qu’on appelle l’instabilité de Grinfeld. Il est vrai qu’une couche mise en contrainte dans la couche (il y a bien une force qui pousse « sur le côté ») tend à plier, ou, pour ainsi dire, préfère plier. J’ai dit plus haut que c’était dû à une répartition plus favorable des forces et des déformations, dans la chose pliée.
Mais la nature peut aboutir à la même forme, sans plier, si elle est capable de réarranger la couche, soit par des déplacements des cellules individuelles, soit par des différentiels de croissance. Ce qui compte c’est d’une part l’état initial : il y a une poussée dans une feuille plate, et d’autre part l’état final : c’est plié ; mais il existe trente-six façons de passer de l’état ini- tial à l’état final. A vrai dire, on ne sait pas exactement comment le tissu vivant s’arrange avec les déformations individuelles, les déplacements individuels et les multiplie-cations cellulaires individuelles. I1 semble bien que le résultat soit une instabilité de pli par différentiel de crois- sance : ça se développe plus ici, moins là, et non directe- ment par pli.
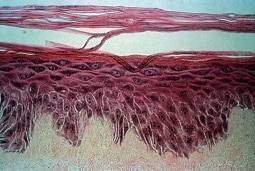
On voit les sillons de la peau, et les cellules formant l’épi- derme. Ces cellules se dessèchent et deviennent plates et dures à la surface ; là, on les distingue à peine. Un peu sous la surface, on voit des petits ronds, les cellules. On remarquera sans peine que la répartition de ces ronds et leur forme ne sont pas homogènes. Dans le fond des plis les cellules sont nombreuses et petites, dans le haut des galbes les cellules sont plus rondes et éparses.
Si le tissu humain était un tissu homogène plié, il n’y aurait pas de différence entre les régions. C’est parce que les cellules répondent aux forces d’étirement/compression, qu’elles se divisent plus dans le fond des plis, où la contrainte mécanique est plus grande. Sous l’effet de cette contrainte et du plus grand nombre de divisions, leur taille est plus petite, et leur densité plus grande, de là elles remontent vers les régions galbées (cet effet est également observé dans le cerveau). Au final, ça a l’air plié, mais ça n’a pas, à proprement parlé, plié. Ce genre de subtilité explique qu’on puisse rencontrer des difficultés, au coup par coup, pour expliquer une observation biologique par de la physique.
Green, par exemple, décrit la formation des méristèmes comme un mécanisme de pli mécanique (un peu comme de la tôle ondulée), mais il ne dit nulle part comment le végétal s’arrange pour rester plié. Or, lorsqu’on plie, mettons, un mètre de menuisier, il reprend sa forme initiale, à moins que quelque chose n’encaisse l’énergie de déformation(comme dans les chocs routiers, où les voitures sont froissées pour de bon). Comment le tissu vivant « absorbe »- t-il les plis pour rester plié ? Green n’en dit rien. Il est plus probable qu’un mécanisme de réarrangement du tissu, plus que de pli, soit à l’œuvre (bien que, dans ce cas aussi, il faille encaisser l’énergie de déformation, au bout du compte).
Vidéo: Où l’on retrouve les empreintes digitales ?
Vidéo démonstrative pour tout savoir sur: Où l’on retrouve les empreintes digitales ?