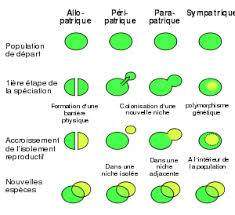
La spéciation sympatrique
 La spéciation sympatrique pose un problème théorique. Pour qu’il y ait spéciation, il faut que des populations divergent au sein d’une espèce, c’est-à-dire s’individualisent génétiquement. Pour cela des différences génétiques apparaissent et les individus s’apparient en fonction d’elles. Il faut donc que ces caractères nouveaux naissent chez plusieurs individus en même temps. A ce premier problème, s’ajoute le fait qu’une divergence entre populations sera favorisée si les hybrides entre ces deux formes sont contre sélectionnés. Mais dans ce cas l’allèle le plus rare est condamné à disparaître et avec lui une des deux populations. Ces problèmes conduisent certains évolutionnistes à douter de la réalité de la spéciation sympatrique. Divers exemples suggèrent que ce type de spéciation est cependant possible en particulier chez les végétaux (voir Briggs & Walters 1997, Ridley 1997).
La spéciation sympatrique pose un problème théorique. Pour qu’il y ait spéciation, il faut que des populations divergent au sein d’une espèce, c’est-à-dire s’individualisent génétiquement. Pour cela des différences génétiques apparaissent et les individus s’apparient en fonction d’elles. Il faut donc que ces caractères nouveaux naissent chez plusieurs individus en même temps. A ce premier problème, s’ajoute le fait qu’une divergence entre populations sera favorisée si les hybrides entre ces deux formes sont contre sélectionnés. Mais dans ce cas l’allèle le plus rare est condamné à disparaître et avec lui une des deux populations. Ces problèmes conduisent certains évolutionnistes à douter de la réalité de la spéciation sympatrique. Divers exemples suggèrent que ce type de spéciation est cependant possible en particulier chez les végétaux (voir Briggs & Walters 1997, Ridley 1997).
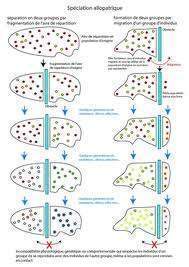
Rhagoletis pomonella est une mouche d’Amérique du Nord dont les larves se nourrissaient initialement des fruits de l’aubépine. En 1864 on découvre des larves qui se développent dans des pommes. Depuis cette nouvelle forme a envahi les vergers et s’est même attaqué à d’autres fruits. Il se trouve que des femelles nées sur des pommes vont préférentiellement pondre dans des pommes et que des mâles nés dans des pommes s’accouplent avec des femelles qui viennent pondre sur des pommes. La durée du développement larvaire est plus courte de 15 à 20 jours dans la pomme que dans les fruits de l’aubépine. Il s’ensuit un isole¬ment reproducteur car les périodes d’activité des adultes des deux formes ne sont pas synchrones. L’analyse électrophorétique confirme cet isolement en montrant qu’elles diffèrent par leurs enzymes. On est bien en face d’un isolement reproducteur et d’une spéciation sympatrique.
Les exemples de spéciation sympatrique sont assez bien documentés chez les plantes avec les spéciations par hybridation. On a ainsi découvert en 1950 aux États-Unis deux nouvelles espèces du genre Tragopogon qui se sont avérées être des hybrides tétraploïdes à 2n = 12 de deux espèces à 2n = 6. Ces nouvelles espèces semblent provenir de plusieurs événements d’hybridation. On a montré que ces hybrides possédaient des formes nouvelles de certaines enzymes grâce à la combinaison de composants des enzymes parentales (travaux de Roose & Gottlieb cités par Ridley 1997). Des espèces d’Iris hybrides diploïdes ont également été découvertes dans les marais de Louisiane. Chez les végétaux la spéciation par hybridation est favorisée par la multiplication végétative. C’est justement le cas des iris qui se multiplient par division de souche par leurs rhizomes. En effet, une fois qu’un hybride se forme (ou plusieurs) la taille de la population peut augmenter assez vite par reproduction asexuée, en particulier si les hybrides sont adaptés à des niches différentes de celles des espèces parentes. C’est souvent le cas car les hybridations se produisent en limite des habitats de ces espèces parentes. Si ce mécanisme permet d’expliquer l’apparition de certaines espèces, il ne suffit pas i expliquer toutes les spéciations car il suppose la préexistence des deux espèces qui s’hybrident qui ont dû apparaître par un autre processus.
La spéciation sympatrique semble plus aisée à concevoir chez les végétaux car, chez ces organismes, la reproduction et le développement diffèrent notablement de ce que l’on rencontre chez les animaux. A cela plusieurs raisons, liées à la biologie des organismes (Niklas .997) :
- le développement embryonnaire est plus simple chez les végétaux que chez les animaux et il est possible que la cohabitation de deux programmes génétiques différents au sein i un hybride soit plus « supportable » dans ce cas. On peut illustrer cette « simplicité » du développement, en comparant le nombre de types cellulaires différents présents dans ces organismes, quelques dizaines chez les Angiospermes, plus de deux cents chez les Mammifères ;
- la mobilité et les perceptions sensorielles des animaux permettent d’établir des barder es à l’hybridation plus efficaces alors que les végétaux s’intercroisent à tout va ; il n’y a pas de contrôle comportemental ;
- de nombreuses plantes ont une croissance indéfinie grâce à des méristèmes, ce qui permet une importante multiplication végétative et donc la production de très nombreux individus susceptibles de constituer une population clonale. Les mutations affectant les méristèmes ou les individus au sein de ces populations engendrent une certaine diversité génétique ;
- les hybrides stériles peuvent devenir fertiles par doublement spontané de leur garniture chromosomique ;
- de nombreuses plantes sont capables d’autofécondation.
L’autopolyploïdie et l’allopolyploïdie sont alors des événe¬ments beaucoup plus fréquents chez les végétaux que chez les animaux et permettent d’interpréter les cas de spéciation présentés dans ce paragraphe ainsi que beaucoup d’autres. À tel point que pour certains botanistes la spéciation par allopolyploïdie serait responsable de la formation de 25 à 50% des espèces actuelles de plantes, ce qui, si l’on pousse le raisonnement à l’extrême en tenant compte de la biomasse que représentent ces organismes, conduit à penser que la spéciation sympatrique est plus fréquente que la spéciation allopatrique si on tient compte des animaux et des végétaux ensembles (Niklas 1997). Sans aller jusqu’à cette extrémité, il faut bien reconnaître que les mécanismes de l’évolution, en grande partie fondés sur la sexualité, sont sensiblement différents selon les modalités de la reproduction des êtres vivants.