Fusions et spéciation stasipatrique
 On connaît chez certaines espèces des populations qui se caractérisent par des réarrangements chromosomiques et en particulier la fusion de certains d’entre eux. Blanc (1982) cite l’exemple étudié par White (1968) de certaines sauterelles australiennes. Les travaux menés à Montpellier au sein de l’institut des Sciences de l’Évolution ont montré l’existence de telles populations chez les souris du genre Mus en Europe.
On connaît chez certaines espèces des populations qui se caractérisent par des réarrangements chromosomiques et en particulier la fusion de certains d’entre eux. Blanc (1982) cite l’exemple étudié par White (1968) de certaines sauterelles australiennes. Les travaux menés à Montpellier au sein de l’institut des Sciences de l’Évolution ont montré l’existence de telles populations chez les souris du genre Mus en Europe.
Le caryotype classique comporte 2n=40 chromosomes, acrocentriques, chez les souris. On connaît plusieurs groupes de populations « robertsoniennes », c’est-à-dire comportant une ou plusieurs fusions dans leur caryotype, un peu partout en Europe. Dans ce type de remaniement, deux chromosomes acrocentriques fusionnent au niveau de leur centromère pour former un chromosome métacentrique. L’étude de la nature et de la distribution de ces fusions montre que chaque groupe, possédant ses propres fusions, a une origine séparée et donc que les fusions peuvent apparaître plusieurs fois au sein des populations. De plus, l’analyse électrophorétique de ces populations montre qu’elles ne forment pas un groupe homogène, mais qu’elles se rattachent génétiquement au groupe géographique auquel elles appartiennent (Britton-Davidian 1985). L’extrême similitude génétique des populations « standards » et des populations « robertsoniennes » dans une région donnée conduit à penser que les populations à caryotype remanié ont une origine très récente (Britton-Davidian et al. 1989). On considère en effet que la divergence génétique qui se met en place dans les populations isolées n’a pas eu le temps de faire son effet.
La thèse de Auffray (1988) présente un des exemples connus de ces populations à chromosomes fusionnés. Dans la plaine du Pô (Italie), dans un rayon de 30 à 100 km autour de la ville de Milan, existent des races chromosomiques de la Souris domestique caractérisées par une réduction du nombre des chromosomes par fusion robertsonienne. Huit populations ont été déterminées dont le nombre de chromosomes varie entre 38 et 22. Les populations distinguées sur la carte et comportant le même nombre de chromosomes diffèrent entre elles par la nature des chromosomes fusionnés. On peut remarquer que ces populations à nombre réduit s’excluent géographiquement les unes les autres et ne cohabitent pas avec les populations « standards » à 40 chromosomes. Elles peuvent s’hybrider entre elles mais si les croisements entre les populations robertsoniennes et les populations à 40 chromosomes donnent des hybrides fertiles, les hybrides obtenus par croisement entre elles des populations à chromosomes fusionnés sont en général stériles par suite de difficultés d’appariement des chromosomes homologues lors de la prophase de la division réductionnelle de la méiose. Seules échappent à cette règle les populations ne différant que par une fusion (Montgellard 1985). De telles populations qui s’excluent (compétition ?) et sont en partie reproductivement isolées posent un problème quant aux mécanismes de leur apparition. Comment concevoir de telles divergences sans phénomène apparent de fractionnement de l’habitat conduisant à des isolats ? Comment des fusions chromosomiques peuvent-elles se répandre au sein des populations ? Il s’agit de plus d’un phénomène récent dans le cas des souris du genre Mus car la colonisation de l’Europe du Sud par cette espèce date du Néolithique .
Ce réarrangement pourrait se répandre dans la population en admettant que les individus porteurs donnent plus de gamètes à chromosomes remaniés que de gamètes normaux. Ces derniers supplanteront les autres si les chromosomes fusionnés à l’état homozygote procurent un avantage sélectif. Mais ce modèle est basé sur de nombreuses suppositions dont certaines n’ont pu être démontrées. En effet, comme l’indique Auffray dans sa thèse, aucun avantage sélectif des différentes fusions chromosomiques n’a été mis en évidence et les études menées au niveau gamétique à la méiose ont montré soit la transmission des deux formes avec la même probabilité, soit la transmission préférentielle de celle portant les chromosomes non fusionnés. Par contre, il semble bien, dans le cas des souris, que ces fusions touchent essentiellement de petites populations isolées. Les travaux menés en Italie par les chercheurs de l’institut des Sciences de l’Évolution de Montpellier ont montré en effet que les populations à nombre réduit de chromosomes sont toutes des populations commensales de l’Homme et ce fait semble généralisable à l’ensemble des populations robertsoniennes connues en Europe et en Afrique du Nord (Auffray 1988). Ce fait est en accord avec le modèle de spéciation stasi- patrique proposé par White (1968) dans la mesure où ces populations vivent dans des fermes relativement isolées les unes des autres pouvant être colonisées par des groupes réduits d’immigrants.
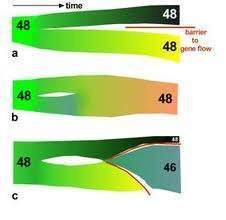
Le modèle défini par White est relativement simple dans son principe. Pour lui, une fusion serait apparue une seule fois chez un individu dans une petite population isolée. Sous l’effet de la dérive méiotique et de la consanguinité le métacentrique passe à l’état homozygote. Plus l’effectif de la population est petit, plus la fixation est rapide. Les hybrides hétéro-zygotes étant moins fertiles du fait des problèmes d’appariements des homologues à la méiose, l’isolement des homozygotes devient plus marqué et les flux géniques avec la population d’origine se réduisent. 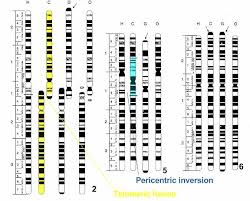 L’adaptation aux conditions écologiques locales se met alors en place. Cela aboutit à l’occupation d’une nouvelle niche écologique et la nouvelle population peut coloniser l’aire de répartition de la population originelle . Ce processus est encore accentué par la fixation de nouvelles fusions. Pour Baker & Bickham (1986), le modèle apparaît applicable à de nombreuses espèces de Mammifères, réparties dans différents ordres, essentiellement les Insectivores et les Rongeurs. Néanmoins, si ce modèle est convaincant en ce qui concerne l’efficacité des barrières d’isolement reproducteur entre espèces à caryotypes remaniés, il reste de nombreuses zones d’ombre concernant l’apparition des fusions à l’état homozygote et leur diffusion dans les populations.
L’adaptation aux conditions écologiques locales se met alors en place. Cela aboutit à l’occupation d’une nouvelle niche écologique et la nouvelle population peut coloniser l’aire de répartition de la population originelle . Ce processus est encore accentué par la fixation de nouvelles fusions. Pour Baker & Bickham (1986), le modèle apparaît applicable à de nombreuses espèces de Mammifères, réparties dans différents ordres, essentiellement les Insectivores et les Rongeurs. Néanmoins, si ce modèle est convaincant en ce qui concerne l’efficacité des barrières d’isolement reproducteur entre espèces à caryotypes remaniés, il reste de nombreuses zones d’ombre concernant l’apparition des fusions à l’état homozygote et leur diffusion dans les populations.