La spéciation géographique : Des spéciations par fractionnement et par colonisations
 Un cas classique est celui proposé pour les goélands eurasiatiques. Il n’a cependant plus, à l’heure actuelle, qu’une valeur historique. On le trouvera dans Mayr (1974) et Blanc (1990) et il est repris dans la plupart des ouvrages consacrés à l’évolution. Les aires de répartition des différents taxons ont changé et la systématique du groupe est en constant remaniement. Chez ces goélands, on distingue à l’heure actuelle :
Un cas classique est celui proposé pour les goélands eurasiatiques. Il n’a cependant plus, à l’heure actuelle, qu’une valeur historique. On le trouvera dans Mayr (1974) et Blanc (1990) et il est repris dans la plupart des ouvrages consacrés à l’évolution. Les aires de répartition des différents taxons ont changé et la systématique du groupe est en constant remaniement. Chez ces goélands, on distingue à l’heure actuelle :
- Larus argentatus à pattes roses de l’Amérique du Nord (sous-espèce smithsonianus), de l’Europe du Nord (sous-espèce argentatus en Scandinavie et Danemark et argenteus sur les berges de la Mer du Nord, dans les îles Britanniques, en France et en Islande) et du nord-est sibérien (sous-espèce vegae) ;
- Larus cachinans à dos gris et pattes jaunes répandu de la Méditerranée à la Mongolie avec quatre sous-espèces reconnues qui sont d’ouest en est : atlantis (Canaries, Açores et Madère), michahellis, cachinans et mongolicus auxquelles il faut rajouter les populations du pourtour de la baltique qui constituent la sous-espèce omissus, le statut des populations du littoral atlantique de la péninsule ibérique est discuté car ces oiseaux, qui différent de ceux de Méditerranée par la taille et la couleur du manteau et qui présentent des ressemblances avec argentatus, sont parfois placés dans la sous-espèce atlantis ;
- Larus glaucoides à dos gris et pattes roses de l’Arctique nord américain avec trois sous-espèces.
- Larus fuscus qui comprend plusieurs sous-espèces du nord de l’Europe à la Sibérie (d’ouest en est : graellsii, fuscus, heuglini, taimyrensis), les aires de distribution des deux sous-espèces européennes (graellsii et fuscus) relèvent sans doute plutôt d’un cline, le statut taxinomique des sous-espèces sibériennes est discuté car certains auteurs en font des sous- espèces de fuscus, d’autres d ‘argentatus, d’autres regroupent heuglini, taimyrensis mais aussi argentatus vegae dans l’espèce heuglini (voir Stepanyan 1990 cité par Filchagov et al. 1992) ;
- Larus armenicus niche dans l’intérieur des terres en Asie Mineure.
On peut remarquer :
- que ces espèces sont en général allopatriques, ou tout au moins l’étaient encore récemment, sauf argentatus avec glaucoides en Amérique du Nord et avec fuscus en Europe ;
- qu’elles sont subdivisées en un certain nombre de sous-espèces dont les statuts taxonomiques sont encore discutés, par exemple, l’absence de couples mixtes dans la zone de contact entre Larus cachinans cachinans et L. c. michahellis suggère qu’il s’agit en fait de deux espèces (Isenmann com. pers.) ;
- qu’il existe des zones d’hybridation au contact entre les espèces allopatriques ;
- que des couples mixtes apparaissent parfois entre les espèces sympatriques, c’est le cas entre argentatus et fuscus avec un couple mixte fertile qui a donné trois jeunes sur le littoral atlantique français et des individus hybrides qui ont été identifiés en Bretagne (Yésou 1991);
- que les passages entre deux sous-espèces se font parfois graduellement, c’est le cas entre fuscus graellsii et fuscus fuscus où le passage se fait par la forme intermediaris ;
- que les sous-espèces sibériennes de fuscus ont parfois été considérées comme des sous-espèces d’argentatus.
Comment interpréter ces distributions ? Mayr (1974) fait intervenir un fractionnement de l’aire de répartition d’une espèce ancestrale de goéland intervenu au moment des glaciations et qui a conduit à l’isolement de populations dans des zones refuge. Par la suite, avec le recul des glaciers, ces populations ont pu recoloniser les zones plus nordiques jusqu’à entrer en contact avec les autres populations isolées dans les autres refuges. Le long de ces zones de contact, les cas d’hybridation observés montrent que le processus de spéciation, s’il a été réellement engagé, n’est pas achevé. Ceci est confirmé par l’exclusion géographique généralement rencontrée entre ces espèces, exclusion qui semble montrer que la divergence n’est pas allée jusqu’au changement de niche écologique.
On suppose par contre que l’espèce argentatus a colonisé l’Amérique du Nord à partir des populations du nord sibérien (sous-espèce vegae). Ces dernières ont aussi donné naissance à l’espèce fuscus vers l’ouest. Argentatus a ensuite colonisé l’Europe du Nord depuis l’Amérique. Il s’est trouvé là en sympatrie avec l’espèce fuscus avec laquelle il cohabite avec quelques cas d’hybridation. Dans ce cas, la divergence a été suffisamment importante pour permettre une situation sympatrique : les niches écologiques de ces espèces sont différentes (voir Géroudet 1982 et Pons in Yeatman-Berthelot & Jarry 1994 pour les caractères écologiques des deux espèces). Sur les sites de nidification, le goéland brun installe son nid sur des surfaces planes et herbeuses alors que le goéland argenté préfère les substrats rocheux. Migrateur partiel, il pond une dizaine de jours plus tard que le goéland argenté qui est plutôt sédentaire. Le goéland brun est plus piscivore, il semble plus habile pour récupérer les déchets de poissons rejetés par les bateaux de pêche et il se nourrit moins sur les décharges que le goéland argenté qui s’est fait une spécialité de rechercher sa nourriture parmi les ordures ménagères. Des mécanismes comportementaux ont été décrits qui limitent la formation de couples mixtes.
On a donc une ceinture d’espèces issue d’une colonisation circulaire des régions circumpolaires. Les populations de l’extrémité de la chaîne se trouvent en contact avec celles du début de la chaîne sans qu’il y ait hybridation. Notons que, là aussi, on a l’impression que la compétition entre des espèces à l’écologie semblable entraîne une ségrégation spatiale et que seules les espèces dont les niches diffèrent peuvent cohabiter. Nous discuterons plus ample¬ment par la suite de la réalité de ce phénomène de compétition.
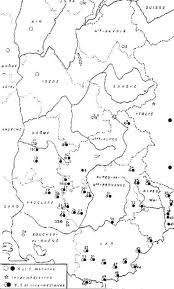
Ce modèle pédagogique (trop ?) doit cependant être pris avec précaution comme le montrent les difficultés de la systématique des goélands, les conséquences des changements d’aire de répartition de certaines de ces espèces (Figure 1.11, voir Yésou 1991, Yeatman- Berthelot & Jany 1994) et des études génétiques récentes. À l’heure actuelle cachinans colo¬nise peu à peu le littoral atlantique français vers le nord à partir des populations de Méditerranée et il a atteint le sud de la Bretagne. Dans le même temps, fuscus a étendu son aire de répartition vers le sud, colonisant le nord-ouest de l’Espagne et du Portugal (premier cas en 1973) et la France au sud de la Bretagne (à partir de 1980). Une colonie s’est même installée en 1983 dans le delta de l’Ebre sur le littoral méditerranéen espagnol. Argentatus pour sa part a progressé vers le sud à partir de la Bretagne, atteignant le Bassin d’Arcachon en 1989. Il existe donc de nos jours une large zone de sympatrie où cohabitent les espèces argentatus, fuscus et cachinans sur le littoral atlantique français et une zone de sympatrie entre fuscus et cachinans en Espagne et au Portugal. Des couples mixtes argentatus X fuscus mais surtout fuscus X cachinans y sont observées. C’est le cas en particulier en Espagne où se reproduisent 350 à 400 couples de Larus fuscus avec 30 à 50 couples mixtes fuscus X cachinans (S.E.O./Birdlife 1997). Par contre, aucun couple mixte entre argentatus et cachinans n’a été observé à ce jour. Or ces deux goélands ont été pendant longtemps considérés comme deux sous-espèces de la même espèce et ils se ressemblent en apparence beaucoup plus que fuscus et cachinans. Les phylogénies de goéland basées sur les couleurs du plumage et des pattes sont donc sans doute à manier avec beaucoup de précaution. Larus fuscus, malgré son dos brun, serait plus proche de Larus cachinans au dos gris que de Larus argentatus lui aussi au dos gris. Enfin, des études génétiques récentes (Crochet non publié) basées sur l’analyse de l’ADN mitochondrial n’ont pas montré une parenté particulière entre les goélands argentés européens (<argentatus argentatus et argentatus argenteus) et nord américains (argentatus smithsonianus).
Les sapins du genre Abies et le complexe d’espèces du groupe des Pins noirs (Pinus nigra au sens large) permettent d’illustrer la coexistence de ces deux types de spéciations chez les végétaux . Chez les sapins, il existe trois espèces à grandeaire de répartition : Abies alba en Europe, Abies sibirica en Asie septentrionale et Abies bal- samea en Amérique du Nord. La région méditerranéenne se singularise quant à elle par l’existence de dix espèces pratiquement toutes allopatriques. La côte ouest américaine est pour sa part riche de vingt espèces environ et quarante espèces se répartissent entre la chaîne de l’Himalaya, les montagnes chinoises et celles du Japon. C’est donc en limite sud de l’aire de répartition du genre, là où les habitats favorables sont morcelés et limités à certains reliefs, ce qui isole des populations, effet d’isolement accentué dans le bassin méditerranéen par les multiples îles, que le nombre d’espèces est le plus élevé. On peut supposer que lors du dernier réchauffement climatique post glaciaire, des populations de sapins se sont retrouvées isolées dans les zones montagneuses, donnant ainsi naissance à ces nouvelles espèces. D’autres sont apparues par colonisation aléatoire d’îles mais là se pose le problème de la dissémination à longue distance de ces végétaux. Les trois espèces à grande aire de répartition occupent des zones favorables homogènes sur de grandes surfaces, sans barrière aux échanges de gènes et sur lesquelles aucune spéciation n’a donc pu se réaliser. On peut expliquer de même l’existence des quatre espèces totalisant quinze sous-espèces de pins noirs du bassin méditerranéen par les nombreuses conditions d’isolement créées par la complexité du relief et le grand nombre d’îles de cette région du monde. On notera que parmi les sittelles, qui sont des petits passereaux forestiers, une espèce la Sittelle corse Sitta whiteheadi est inféodée à Pinus laricio poiretana. C’est un exemple de parallélisme entre une spéciation chez les animaux et une spéciation chez les végétaux. Il faudrait vérifier si la Sittelle de Krüper Sitta krueperi est également associée à Pinus nigra pallasiana en Turquie. La Sittelle kabyle Sitta leudanti dAlgérie n’est par contre pas limitée à Abies numidica comme les premières publications l’avaient affirmé. On la rencontre largement dans les chênaies à Quercus canariensis, Q. afares ou Q. suber (Bellatreche 1991, Isenmann com. pers.). Cependant c’est malgré tout dans des massifs isolés que cette sittelle se rencontre et on peut considérer que la spéciation est liée à l’insularité.