Le modèle de Turing:pourquoi le léopard a-t-il des taches?
Alan Turing
De son vivant. Alan Turing est l’auteur d’un unique article de biologie théorique, paru en 1952, mais il est magistral. Il présente un nouveau modèle de formation de structures par « réaction-diffusion » connu simplement sous le nom de « modèle de Turing ». Le titre de cet article est « Base chimique de la morphogenèse ». Son dernier texte, posthume, s’intitule, lui : « Description succincte de la croissance des marguerites ». Comment Alan Turing espérait-il expliquer la formation des plantes ? Son premier souci était d’expliquer l’apparition de quelque chose à partir de rien, c’est-à-dire l’apparition de choses non uniformes comme des creux et des bosses, des taches blanches ou noires, non pas à partir de strictement rien, mais à partir d’un milieu uniforme : une mer d’huile, un monochrome. C’est une question profonde, qui concerne de nombreux domaines.
Par exemple : l’œuf de poule paraît à peu près homogène quand on le mange dans le coquetier, or il peut produire un poussin en quelques semaines (s’il est fécondé et couvé, ce qui n’est évidemment pas le cas de l’œuf dans votre réfrigérateur). Ce problème d’embryologie est fort complexe, il va sans dire. Quelle est la nature même de cette complexité ?
On peut attaquer ou réduire la difficulté en commençant par l’organisation des rosettes au centre des fleurs, voilà qui est sans doute plus simple qu’un bébé entier. Mais parmi ces structures botaniques, les plus simples se résument à la succession droite gauche des feuilles de certaines plantes. De fil en aiguille la question se réduit à : comment faire apparaître des petits domaines, bosses, pointes, bourgeons dans un milieu « plat » ? On verra plus tard comment faire des oreilles ou des museaux, des ailes de papillon ou des râbles de lapin.
L’idée de départ de Turing était que tout cela venait de la chimie, mais une chimie subtile, dont il est l’inventeur. Comme nous allons le voir, il s’agit d’une chimie de compétition. Lui-même appelait sa théorie de l’embryologie chimique. Pour fabriquer un embryon, expose-t-il, il faut que des concentrations en espèces chimiques s’organisent toutes seules pour produire un embryon : un peu plus de ceci par ici, un peu plus de cela par là, et ça finira par faire des choses inhomogènes, peut-être un doigt ou un pied.
Mais il faut que cette inhomogénéité vienne d’elle-même « à partir de rien », pour ainsi dire, de l’intérieur du système. Pour commencer, soyons un peu moins ambitieux. Peut-on faire de la chimie qui produise de simples taches, ou quelques zébrures, des peaux de girafe ou de zébu, à partir de rien ?
A partir de strictement rien, non. Pas possible. Mais la réponse est : à partir de très, très peu, c’est possible. Certains systèmes sont instables, en sorte qu’ils accroissent les petites irrégularités locales, au point qu’à la fin, en lieu et place de l’uniformité, on a quelque chose de plus compliqué, de plus divers, en un mot : il se passe des choses intéressantes, qui se révèlent d’elles-mêmes à partir d’un milieu presque complètement uni, dont les inhomogénéités ou grumeaux sont, au départ, imperceptibles à l’œil nu.
Je précise ici que les chercheurs contemporains croient souvent que l’exemple des taches de léopard et des peaux de girafe, ou l’exemple des marguerites, est venu bien après, que ce sont des inventions, des applications, postérieures au modèle de Turing proprement dit. Il n’en est rien : Alan Turing a exposé explicitement l’application de ses modèles à des taches de léopard ou de girafe, et à la rosette des marguerites. Ainsi, ce qui intéressait Turing, c’était de produire des petits domaines, des petites taches de concentration, taches de léopard, ou zones de formation d’ébauches de feuilles, grains de beauté ou racines de poils, cartilages des doigts ou distribution des pores de la peau.
Aujourd’hui, les théoriciens ont appliqué les idées de Turing à la plupart des systèmes où apparaissent des structures à peu près régulières : points, lignes, bandes zébrées ou en zigzag, carrés ou hexagones. On aurait envie de dire « bien au- delà, sans doute, de ce qu’il avait imaginé », mais l’honnêteté oblige à dire « non, pas tellement au-delà de ce qu’il avait proposé ». Dans le modèle et les idées de Turing, il y avait déjà tout ce qu’on voit aujourd’hui, ou peu s’en faut, et lorsqu’on songe au caractère primitif de son ordinateur, en comparaison des nôtres, on se dit que l’essentiel était dans la puissance de son esprit, et peu de chose dans la machine.
Mais expliquons comment marche le mécanisme de Turing. C’est une affaire de compétition, nous l’avons évoqué plus haut. Il y a deux équipes qui ne jouent pas le même jeu, mettons, les Bleus et les Rouges. Les Bleus tapent sur la tête des Rouges, pour les écraser. Les Rouges, eux, sont moins sévères : ils ne font pas grand-chose, sauf se reproduire entre eux, et, parfois, devenir Bleus. Le nombre naturel de Rouges tend donc à s’accroître. Là où les Rouges s’accroissent, les Bleus s’accroissent aussi, en principe. Mais les Bleus ont une particularité, ils se dispersent plus vite que les Rouges.
Il y a donc une tendance générale à l’augmentation des Rouges, s’ils étaient tout seuls, et une tendance générale à la dilution des Bleus, s’ils étaient tout seuls. Mais voilà : les Bleus ne sont pas tout seuls, et les Rouges non plus. On met les deux ensemble, et les Bleus s’en prennent aux Rouges. Supposons qu’on parte d’une situation initiale où il y a et des Bleus et des Rouges partout (disons, c’est à peu près uniformément marron), est-ce que ça va rester comme ça éternellement ? Est-ce que ça va devenir tout Rouge, ou tout Bleu, ou bien obtiendra-t-on un mélange de Rouge et de Bleu ? Eh bien la réponse est : ça dépend. Ça dépend de la vitesse à laquelle le Rouge et le Bleu se dispersent, une fois qu’ils sont produits. Supposons que le Rouge se disperse peu : là où il s’en produit, il s’en produit de plus en plus (effet de croissance exponentielle, comme pour une population), et ça a toutes les chances d’augmenter beaucoup.
Comme le Rouge augmente beaucoup, il produit de plus en plus de Bleu, mais comme le Bleu se disperse, il a du mal à freiner sur place l’augmentation du Rouge, qui a pris trop d’avance. Mais le Bleu se disperse vite autour du Rouge, qui, lui, ne se disperse pas très vite. Résultat : il n’y a pas assez de Bleu localement pour écraser le Rouge, mais un peu plus loin, il y a moins de Rouge et encore assez de Bleu : là, le Bleu est assez fort pour écraser le Rouge. Par conséquent, si une tache de rouge se forme quelque part, elle sera entourée, non loin de là, par une zone de Bleu. On peut répéter le raisonnement sur une distribution de taches de Rouge, et montrer ainsi qu’on peut obtenir un ensemble de taches de Rouge, cernées par du Bleu. Compte tenu de sa faible dispersion, le Rouge reste, en un certain point, assez fort, mais autour de lui ses défenses sont trop faibles et il laisse le Bleu l’emporter.
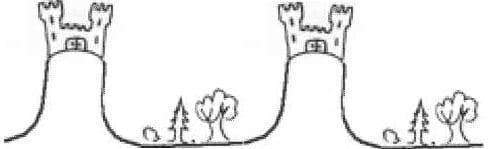
C’est un peu le principe du château fort : les villageois concentrent leurs forces en un point, imprenable, et laissent les brigands s’emparer des forêts autour. Mais comme les brigands n’ont pas de chef et qu’ils se promènent partout en bande désorganisée, il n’y en a jamais assez pour conquérir tous les châteaux forts. Résultat : il y a une distribution de châteaux forts robustes, baignant dans un océan de zones peu sûres. Même si certains villageois deviennent brigands, le nombre de villageois augmente près du château, qui reste imprenable, tandis que les brigands détroussent et font fuir tous les villageois égarés dans la forêt.
En termes scientifiques, on appelle les habitants du château les activateurs (ils s’activent eux-mêmes pour être plus nombreux, et de temps en temps ils deviennent brigands), et les brigands les inhibiteurs (ils font ce qu’ils peuvent pour tout détruire, empêcher l’érection de nouveaux châteaux). Suivant les valeurs des conditions initiales, et suivant la taille du système, on peut n’avoir que des villageois, que des brigands ou bien un mélange ordonné de brigands et de villageois.
Bon, j’avoue qu’il n’est pas facile de décrire de façon un peu originale le modèle de Turing : tout a été dit. Les requins et les crevettes, les moutons et le raigras, les noirs et les blancs, la gauche et la droite, le virus et l’antibiotique, les bons et les méchants, le renard et le corbeau, tout a été ou sera utilisé comme métaphore pour décrire la formation de taches par le modèle de Turing, qu’on appelle aussi modèle activateur-inhibiteur. Dans un article de vulgarisation très complet, James Murray, un mathématicien anglais, choisit l’exemple des feux de forêts et des pompiers.
Ce modèle est relativement simple, et il donne des distributions de taches de léopard ou de girafe « très raisonnables ] », suivant le mot de Turing à un de ses collègues, dans une lettre. Si l’on effectue une simulation numérique sur un pays carré, on obtient des taches qui occupent régulièrement l’espace, comme un damier. Si on effectue la simulation sur un domaine en forme de peau d’animal, on obtient des taches évoquant les zèbres, les girafes, les léopards, et ainsi de suite.
Pour expliquer la variété des taches des animaux, on pourra considérer des variétés de forme de peau, ou bien des variétés de date d’apparition des taches. En effet, ce qui caractérise, au final, la forme des taches, est une longueur ou une épaisseur caractéristique. C’est la taille d’un domaine de couleur unie : un ocelle, une tache de girafe, la largeur d’une zébrure. Mais comment expliquer alors que certains animaux n’ont pas de taches, mais sont plutôt à moitié noirs et à moitié blancs ? Murray a beaucoup étudié ces questions, et pense expliquer les variétés de taches d’animaux par un modèle de Turing ayant lieu à un âge, ou un autre, de la gestation de l’animal. Suivant que le mécanisme de Turing se met en place quand l’animal (le fœtus) est très grand, in utero, ou bien très petit, on pourra rentrer beaucoup de taches dans la surface de la peau, ou bien seulement une demi-tache, et alors l’animal sera à moitié noir et à moitié blanc.
Un autre exemple est celui de la queue des léopards. Selon Murray, le modèle de Turing explique parfaitement la variation du dessin sur la queue des félins. On observe sur une vraie queue de léopard que le haut de la queue voit des taches, comme sur le corps de l’animal, que le dessin sur le milieu de la queue est formé plutôt d’anneaux, et qu’au bout, certes plus petit et pointu, la queue est en général de couleur unie. Cette succession de motifs s’explique très aisément si l’on introduit un modèle de Turing formant des motifs sur un cône. Tout au bout de la queue, celle-ci est si petite qu’elle est nécessairement tout unie. Tout près du corps, la queue est assez large pour que plusieurs taches apparaissent.
Entre ces deux extrémités, l’uni se décompose en taches, en formant d’abord des anneaux. Tout cela est très séduisant, bien que, à ma connaissance, il n’y ait aucune preuve formelle d’un tel mécanisme dans les animaux. Sans attendre la confirmation des biologistes, on peut dire que le modèle fait qualitativement une prédiction juste : un animal qui a des taches sur le corps peut avoir des zébrures sur la queue, alors que l’inverse n’est pas vrai. Si l’animal est zébré, la queue sera zébrée ou unie, mais elle ne peut pas présenter des taches.
![]()
Le mécanisme de Turing a été étendu aux coquillages, et le lecteur intéressé trouvera de merveilleuses simulations dans un beau livre d,art scientifique, si l’on ose dire : The Algorithme Beauty ofSea Shells de Hans Meinhardt. Ce livre explique comment former des motifs de conques ou d’escargots de mer à l’aide de couples chimiques du type « réaction-diffusion ».
Vidéo: Le modèle de Turing:pourquoi le léopard a-t-il des taches?
Vidéo démonstrative pour tout savoir sur: Le modèle de Turing:pourquoi le léopard a-t-il des taches?