
Le modèle neutraliste de l'évolution moléculaire : La reconstitution de phylogénies
écrit le: 27 juin 2013 par admin
La reconstitution de phylogénies
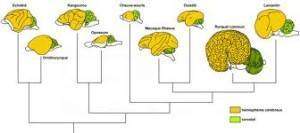
Dans un premier temps, l’étude des substitutions d’acides aminés dans les enzymes sériques d’espèces apparentées, permet, dans les cas bien documentés, de tester la validité du modèle de l’horloge moléculaire. C’est le cas, par exemple, pour les drosophiles des îles Hawaii. Cet exemple est particulièrement intéressant car les différentes espèces sont confinées à des volcans apparus successivement selon un axe nord-ouest/sud-est et datés très précisément par la méthode potassium-argon. On s’intéresse à un groupe d’espèces, planitibia, constitué de mouches de grande taille peuplant les forêts humides d’altitude et dont la vitesse de reproduction est assez lente, deux générations par an au maximum (Carson 1976), supposée être la plus récente du groupe, et en abscisse l’âge de la plus vielle roche émergente de chaque volcan. La distance génétique est en fait calculée à partir de l’estimation de l’identité génétique des populations, c’est-à-dire du calcul des probabilités pour que des gènes homologues portent les mêmes allèles avec les mêmes fréquences dans les populations comparées. Le détail de ces calculs est exposé dans Lucotte (1983) et Kimura (1990). La droite formée par l’alignement des points montre que les divergences génétiques s’accumulent à un taux de 1% tous les 20 000 ans. Les estimations fournissant une fourchette de vitesse de divergence comprise entre 1% pour 25 000 ans et 1% pour 15 000 ans sont moins satisfaisantes. Selon cette figure, on a bien l’impression d’une évolution à vitesse constante au plan moléculaire.
Un autre exemple que l’on peut donner de l’utilité de ces comparaisons de séquences consiste à étudier un groupe de protéines que l’on peut qualifier d’aquaporines (Chrispeels & Maurel 1994). Il s’agit de molécules homologues impliquées dans les échanges membranaires et le contrôle du potentiel hydrique cellulaire. Ces protéines sont constituées de deux sous- unités comportant chacune trois domaines nitra-membranaires en hélice. La similitude entre les deux sous-unités est telle que l’on pense que le gène actuel résulte de la duplication d’un gène ancestral . . On a comparé les séquences de six membres de cette famille protéique : MIP, extraite des fibres du cristallin de bœuf ; NOD, extraite des nodules racinaires de soja ; GLP provenant de la membrane de Escherichia coli ; TIP, provenant du tonoplaste de nombreuses plan¬tes supérieures ; BIB, extraite du système nerveux de la drosophile ; GLY, issue de la membrane de Streptomyces coelicolor (Pao et al 1991). Les protéines ont été séquencées et ces séquences sont ensuite alignées grâce à un programme informatique qui permet de détecter le maximum de similitude. Les taux de divergence sont ensuite estimés à l’aide de ce même programme, ce qui permet alors de construire l’arbre phylogénétique le plus parcimonieux.
Le premier enseignement que l’on tire de la comparaison de ces différentes séquences est que les procaryotes sont plus éloignés de tous les autres organismes que ces derniers ne le sont entre eux . Ce résultat n’est guère surprenant et ne justifie pas à lui seul l’utilisation de méthodes aussi lourdes.
Mais il y a mieux. Lorsque l’on compare non plus les protéines entières mais les sous- unités, on constate que chacune forme une entité évolutive. C’est-à-dire qu’il y a plus de similitudes, par exemple, entre les sous-unités n°l du soja et du bœuf (NOD1 et MIP1) qu’entre les deux sous-unités de chaque molécule , soja d’un coté (NOD1 et NOD2) et bœuf de l’autre (MIP1 et MIP2). Cette constatation est même valable lorsque l’on compare Procaryotes et Eucaryotes.
La conclusion de ces observations est que la duplication intragénique, qui a donné naissance à une molécule à six domaines transmembranaires, s’est produite avant la divergence conduisant aux Procaryotes et aux Eucaryotes. On peut ici proposer une datation relative d’un événement génétique important sans avoir recours aux archives fossiles.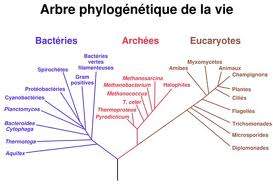
Comme on le voit, la Théorie Neutraliste de l’Évolution Moléculaire explique de manière satisfaisante ou presque un certain nombre de faits concernant la variabilité génétique des populations, la divergence interspécifique et les parentés moléculaires. Elle doit être considérée comme un modèle permettant d’étayer les modèles évolutionnistes classiques. Mais cette théorie ne propose pas de mécanisme de spéciation et pour ce qui est de la compréhension des problèmes de l’Evolution des espèces, elle reste donc incomplète.
Vidéo: Le modèle neutraliste de l’évolution moléculaire : La reconstitution de phylogénies
Vidéo démonstrative pour tout savoir sur: Le modèle neutraliste de l’évolution moléculaire : La reconstitution de phylogénies