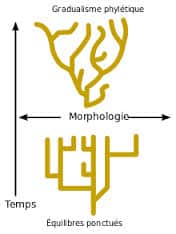
Le saltationisme ou théorie des équilibres ponctués
 L’origine des espèces Charles Darwin, constatant l’absence de formes intermédiaires dans les séries fossiles, reconnaît que là réside l’objection la plus sérieuse que l’on puisse faire à sa théorie. Ce problème est d’ailleurs toujours d’actualité comme le montrait Langaney dans un article de 1997 de la revue La Recherche. En effet, les formations géologiques ne livrent pas au paléontologue des successions de formes manifestement dérivées les unes des autres et illustrant ainsi la transformation progressive des caractères sous l’effet de la sélection ou du hasard. Elles fournissent au contraire des espèces qui, en général, apparaissent brusquement, restent inchangées pendant quelques millions d’années, puis disparaissent sans véritable transition avec celles qui leur succèdent. L’apparition simultanée de tous les plans d’organisation des animaux actuels lors de l’explosion cambrienne il y a 550 millions d’années est un exemple spectaculaire de ces apparitions rapides de formes nouvelles (voir Gould 1991). Ce constat est surtout troublant quand on s’intéresse à la macroévolution, c’est-à-dire à l’apparition des taxons de niveau supérieur à celui de l’espèce,
L’origine des espèces Charles Darwin, constatant l’absence de formes intermédiaires dans les séries fossiles, reconnaît que là réside l’objection la plus sérieuse que l’on puisse faire à sa théorie. Ce problème est d’ailleurs toujours d’actualité comme le montrait Langaney dans un article de 1997 de la revue La Recherche. En effet, les formations géologiques ne livrent pas au paléontologue des successions de formes manifestement dérivées les unes des autres et illustrant ainsi la transformation progressive des caractères sous l’effet de la sélection ou du hasard. Elles fournissent au contraire des espèces qui, en général, apparaissent brusquement, restent inchangées pendant quelques millions d’années, puis disparaissent sans véritable transition avec celles qui leur succèdent. L’apparition simultanée de tous les plans d’organisation des animaux actuels lors de l’explosion cambrienne il y a 550 millions d’années est un exemple spectaculaire de ces apparitions rapides de formes nouvelles (voir Gould 1991). Ce constat est surtout troublant quand on s’intéresse à la macroévolution, c’est-à-dire à l’apparition des taxons de niveau supérieur à celui de l’espèce,
où les formes intermédiaires manquent encore plus cruellement. En effet, si l’évolution est un processus lent, la probabilité de rencontrer des formes intermédiaires entre des classes par exemple doit être plus élevée que la probabilité de rencontrer des formes intermédiaires entre espèces. Or c’est l’inverse que l’on constate et des auteurs comme Denton (1988) trouvent là des arguments à leurs critiques à l’encontre de la théorie de l’évolution. Darwin avançait plusieurs hypothèses pour expliquer cette faiblesse de sa théorie. Il constatait par exemple que seule une faible partie des séries sédimentaires avait été analysée à la surface du globe. La fossilisation lui semblait un phénomène suffisamment exceptionnel pour expliquer ces lacunes, avec en particulier toutes les parties molles qui ne se conservent pratiquement jamais et la possibilité pour les eaux pluviales acides de dissoudre les fossiles. Mais surtout il pensait que l’absence des formes intermédiaires provenait du caractère discontinu des séries sédimentaires. Ces discontinuités ont pour origine l’absence de sédimentation à certaines périodes ou l’érosion ultérieure des dépôts. Cet argument est pertinent quand on constate de nos jours que les discontinuités qui permettent de délimiter des étages représentent parfois une durée supérieure à celle des étages eux-mêmes ou que la différence d’épaisseur des dépôts entre des localités différentes durant le même intervalle de temps peut être considérable. Ainsi, la limite Lutétien-Stampien représente 150 m de dépôts hétérogènes subdivisables en 18 formations dans le Bassin Parisien alors qu’à Biarritz elle se présente sous la forme de 1 500 m de dépôts de calcaires et de marnes uniformes (Pomerol et al. 1987).
Partant du constat que les espèces semblent passer par de longues périodes d’équilibre, appelées stases par ces auteurs, séparées par des transitions rapides ou ponctuations, Niles Eldredge et Stephen Jay Gould proposèrent en 1972 le modèle dit des équilibres ponctués. Pour ces chercheurs, les formes de transition sont absentes des séries fossiles car elles sont peu nombreuses. Elles se forment dans de petites populations isolées en périphérie de l’aire de répartition de l’espèce au sein desquelles les phénomènes de dérive autorisent des vitesses d’évolution élevées. L’espèce en voie de formation sera ainsi fossilisée séparément de l’espèce ancestrale ou pas du tout fossilisée vue la faiblesse de ses effectifs. La nouvelle espèce pourra envahir l’aire de répartition de l’espèce ancestrale dans deux cas : soit en l’éliminant si elle a acquis un avantage compétitif par rapport à elle, soit en cohabitant avec elle si elle a conquis une nouvelle niche écologique. La théorie des équilibres ponctués est basée sur des modèles de spéciation allopatrique. La particularité vient de la vitesse des processus évolutifs qui est accélérée au moment des spéciations. Darwin émettait d’ailleurs cette hypothèse dans son ouvrage quand il écrivait que c’était peut-être des populations locales de l’espèce, de répartition limitée, qui avaient donné naissance aux nouvelles espèces au cours de processus d’évolution accélérés.
Comme d’autres approches, la théorie des équilibres ponctués permet d’expliquer certains aspects de l’évolution. Elle ne les explique cependant pas tous et en particulier des cas d’évolution rapide ont aussi été rapportés dans la partie centrale de l’aire de répartition de certaines espèces et dans de grandes populations. L’apparition de résistance aux insecticides (Poirié & Pasteur 1991) ou l’évolution de la Phalène du bouleau en sont des exemples.
Diverses hypothèses furent avancées pour expliquer les phases d’évolution rapide comme des mutations sur les gènes qui contrôlent le développement. Des mutations sur des protéines de choc thermique (HSP pour heat shock protein) pourraient également expliquer les phases d’évolution rapide. Ces protéines, découvertes en 1973, sont aussi appelées chaperons (ou chaperonines). Elles se lient à d’autres protéines et les aident ainsi soit à acquérir leur structure tertiaire ou quaternaire définitive lors de leur maturation, soit à retrouver cette structure en cas de dénaturation. Pour cela elles se fixent aux résidus hydrophobes des protéines et elles ralentissent le passage à la structure fonctionnelle. Elles empêchent de la sorte que ne s’établissent des liaisons avec d’autres molécules ce qui perturberait l’acquisition de la structure définitive. Contrairement à beaucoup de chaperons qui ne sont exprimées qu’en cas de stress, une protéine de choc thermique, HSP90, est toujours présente dans la cellule eucaryote. Rutherford et Lindquist (1998, voir aussi Grop 1999) ont montré que les descendants de drosophiles mutées sur l’une des deux copies du gène codant pour la HSP90 présentaient des malformations variées en proportion élevée. Des inhibitions chimiques ou thermiques provoquées de cette protéine chez des drosophiles saines ont fait apparaître ces mêmes malformations dans leur descendance. Il semble que la HSP90 supprime, pendant l’embryogenèse, les effets de mutations présentes dans le génome des drosophiles. Ces mutations seraient ainsi normalement silencieuses. Mais en cas d’inactivation de la HSP90, de mutation sur le gène qui la code ou simplement de diminution de sa concentration, elles s’exprimeraient toutes brutalement. Cette hypothèse apporte une base moléculaire à la théorie des équilibres ponctués. En effet, tant que les condi¬tions environnementales sont stables et favorables à l’espèce, des mutations s’accumulent dans le génome, certaines sont sélectionnées mais la protéine chaperon empêche l’expression d’un grand nombre d’entre elles. Mais en cas de stress pendant la phase embryonnaire dû à une dégradation rapide des conditions environnementales, HSP90 exercerait préférentielle- ment son rôle de chaperon. Elle serait alors essentiellement liée à des protéines endomma¬gées.  Elle ne jouerait plus (ou moins) son rôle de masquage des mutations. Un grand nombre de mutations, et par voie de conséquence une variabilité génétique jusqu’alors masquée, s’exprimeraient alors en même temps, ouvrant la voie à une phase d’évolution rapide.
Elle ne jouerait plus (ou moins) son rôle de masquage des mutations. Un grand nombre de mutations, et par voie de conséquence une variabilité génétique jusqu’alors masquée, s’exprimeraient alors en même temps, ouvrant la voie à une phase d’évolution rapide.