Quand les artères et les veines jouent à saute-mouton
Un autre exemple de morphogenèse où un mécanisme connu peut se révéler caché, occulté par un autre, est celui de la formation de la double vasculature enchevêtrée artères-veines. L’image ci-dessous montre une vasculature de rarrière de l’œil d’un porc.
Cette image a été obtenue par un vétérinaire de l’université de Gand : le Dr Simoens, que je remercie.  Il ne s’agit pas, en fait, de l’image de la vasculature, mais, plus exactement, de l’intérieur de la vasculature. En effet, ce moulage est obtenu en injectant dans les vaisseaux principaux de l’araldite ou du mercox (un plastique), et en laissant diffuser le produit jusqu’aux plus petits vaisseaux. Ensuite, on dissout les chairs, et on regarde ce qui reste (le moulage de l’intérieur des vaisseaux). Ce qui frappe, dans cette image, c’est l’existence d’un arbre artériel et d’un arbre veineux enchevêtrés, mais qui communiquent, nonobstant, par les capillaires. Il peut y avoir des artères passant par-dessus ou par-dessous d’autres artères ou veines, et, réciproquement, des veines passant par-dessus ou par-dessous d’autres veines ou artères. Au final tout ce petit monde se met d’accord pour se connecter via des capillaires (il existe assez peu de shunts directs, on en voit quelques-uns dans des vaisseaux légèrement plus gros que les capillaires, ou bien dans certains tissus spécifiques où les vaisseaux forment des reconnexions en arches). Le transfert de l’oxygène (convoyé par le sang) vers les tissus se fait au niveau des capillaires. Il est donc normal que les artères se rétrécissent de plus en plus, de bifurcation en bifurcation, pour aller jusqu’aux capillaires, et il est également normal que le sang passe en revenant dans des vaisseaux qui se regroupent en veines de plus en plus grosses pour retourner au cœur.
Il ne s’agit pas, en fait, de l’image de la vasculature, mais, plus exactement, de l’intérieur de la vasculature. En effet, ce moulage est obtenu en injectant dans les vaisseaux principaux de l’araldite ou du mercox (un plastique), et en laissant diffuser le produit jusqu’aux plus petits vaisseaux. Ensuite, on dissout les chairs, et on regarde ce qui reste (le moulage de l’intérieur des vaisseaux). Ce qui frappe, dans cette image, c’est l’existence d’un arbre artériel et d’un arbre veineux enchevêtrés, mais qui communiquent, nonobstant, par les capillaires. Il peut y avoir des artères passant par-dessus ou par-dessous d’autres artères ou veines, et, réciproquement, des veines passant par-dessus ou par-dessous d’autres veines ou artères. Au final tout ce petit monde se met d’accord pour se connecter via des capillaires (il existe assez peu de shunts directs, on en voit quelques-uns dans des vaisseaux légèrement plus gros que les capillaires, ou bien dans certains tissus spécifiques où les vaisseaux forment des reconnexions en arches). Le transfert de l’oxygène (convoyé par le sang) vers les tissus se fait au niveau des capillaires. Il est donc normal que les artères se rétrécissent de plus en plus, de bifurcation en bifurcation, pour aller jusqu’aux capillaires, et il est également normal que le sang passe en revenant dans des vaisseaux qui se regroupent en veines de plus en plus grosses pour retourner au cœur.
Ce qui est étrange, c’est ce croisement démentiel de vaisseaux gros et moyens, se connectant sans difficulté par des capillaires. C’est un peu comme si, prenant un pommier dans une main (les artères) et un poirier dans l’autre (les veines), vous enfonciez la ramure de pommier dans la ramure de poirier, et qu’aucune branche de l’un ou de l’autre ne venait à se toucher, tandis que toutes les feuilles de l’un se mettaient en contact avec les feuilles de l’autre. Je pense même que cette image est inférieure à la réalité. Comment imaginer un mécanisme spontané, produisant pareil enchevêtrement ? Cela paraît miraculeux. Évidemment, ce n’est pas ainsi que les vaisseaux se forment. On sait cependant que l’écoulement joue un rôle crucial dans la formation des vaisseaux. Si l’on songe à un mécanisme du type des petits canaux d’érosion décrits pour la plage de Granville, on rencontre deux problèmes. Le premier est qu’il faut trouver l’équivalent du sable et l’équivalent de la mer qui rabote, et le second est que les canaux sur la plage ne forment qu’une seule arborescence de « vaisseaux ». et on ne voit pas comment former deux arborescences croisées, enchevêtrées, sans que tout s’effondre lamentablement. Pourtant, un mécanisme du type « Plat Gousset » est bien à l’œuvre dans la formation des vasculatures enchevêtrées. Le premier problème est le plus aisé à dégonfler : l’équivalent du sable, ou plutôt, des interstices entre le sable, c’est le paquet de capillaires qui préexiste dans le tissu, avant la formation des vaisseaux.
Un tissu non encore développé, et en particulier, n’étant le siège d’aucune circulation sanguine, ne possède que des capillaires ; c’est, à trois dimensions, une sorte de gros plat de nouilles assez régulièrement entrecroisées. On distingue ce type de capillaires entre les branches des gros vaisseaux. Ce réseau de capillaires est, toutes choses égales par ailleurs, l’équivalent de la plage au sable uniforme. Peu à peu, la circulation agit sur les vaisseaux pour les élargir. En élargissant des vaisseaux, elle induit localement des écoulements plus grands, d’où un accroissement de l’effet d’élargissement, etc. L’élargissement des vaisseaux sous l’effet de l’écoulement est l’équivalent du sable emporté qui creuse les rigoles, et la formation spontanée d’arborescences est donc liée au phénomène observé au Plat Gousset, par le fait que le plus grand ruissellement de sang dans les vaisseaux est l’équivalent du débit plus grand dans les plus grands canaux. les vaisseaux réels sont plus allongés, ce qui est aussi le cas des arborescences de la plage. C’est dû essentiellement à un effet de réorganisation, ici un lissage des arbres, à l’intérieur de l’arborescence. Mais il faut maintenant expliquer l’enchevêtrement des artères et des veines. Cet enchevêtrement trouve son origine dans ce qu’on appelle en biologie vasculaire la régression capillaire : à mesure que les vaisseaux principaux émergent et se forment, ils se déconnectent des capillaires. sauf aux extrémités, par où le sang sort et irrigue le lit capillaire situé au bout.
De ce fait, le lit capillaire où passe l’arbre n’est, en fait, pas connecté à l’arbre, on remarque bien que les gros vaisseaux ne sont pas connectés aux capillaires. Si vous regardez les veines sur le dos de votre main, pensez que les gros tuyaux ne sont pas connectés, en fait, à ce dos de la main, ce ne sont pas eux qui l’irriguent. Comment se fait donc la croissance et l’enchevêtrement ? D’un côté, des capillaires s’agrandissent en artères, et se déconnectent, au fur et à mesure, desdits capillaires ; ailleurs, des capillaires se transforment en veines, et se déconnectent également, sauf aux extrémités. Si une veine passe au voisinage d’une artère, il n’y a aucun écoulement direct de l’artère vers la veine, car ni l’une ni l’autre ne sont connectées aux capillaires. Les veines s’agrandissent vers l’avant, sous l’effet du sang qui parvient des artères à travers les capillaires (les veines remontent le courant, comme les canaux d’érosion sur la plage) ; quant aux artères, elles s’agrandissent pour une raison analogue, mais en descendant le courant, car ce qui compte, ce n’est pas que le sang aille dans un sens ou dans l’autre, mais qu’il ait une certaine intensité (un débit suffisamment fort élargit les vaisseaux, peu importe que le débit soit dans un sens, ou dans l’autre). Les veines peuvent bien croiser des artères, l’essentiel des artères est invisible pour les veines (et réciproquement). Seules les extrémités des artères et celles des veines restent connectées au lit capillaire, de sorte que les deux arbres se poursuivent par leurs extrémités, et s’enchevêtrent en une danse presque macabre, si l’on pense à ces vasculatures comme à deux arbres morts enlacés. (Cette image est un peu lugubre, étant entendu que le sang est davantage vecteur de vie). On peut effectuer une simulation numérique qui produit ainsi des vasculatures entrelacées, en effectuant deux croissances de deux arborescences analogues aux canaux du Plat Gousset, mais qui se déconnectent au fur et à mesure des capillaires.Il faut préciser deux choses, dans l’image de gauche, je n’ai représenté qu’un quart des capillaires. Ces capillaires sont l’analogue du sable de la plage, ou plutôt, des interstices entre le sable. La figure suivante, dans laquelle on a retiré les capillaires, montre la vasculature enchevêtrée, l’équivalent d’un moulage des canaux enchevêtrés.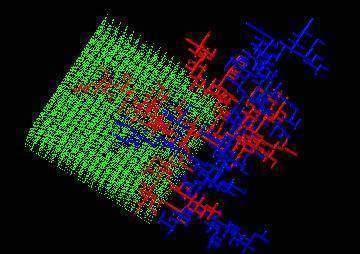
En second lieu, il faut insister sur le fait que, dans cette simulation, tout se passe comme si la mer, au lieu de circuler de la plage vers les canaux, s’écoulait des artères vers les veines, en une marée à sens unique. Dans mon précédent livre, j’ai évoqué le cas de l’embryon de poulet, et j’ai montré qu’une partie au moins de la vasculature de l’œuf était contrôlée par ce mécanisme. Mais la vasculature de l’œuf est une structure bidimensionnelle (ont presque…). J’ai présenté ci- dessus le scénario de construction d’une vasculature tridimensionnelle permettant de fabriquer des vasculatures enchevêtrées. Ce dernier modèle est une conséquence du premier. C’est un développement récent, qui s’est produit entre mon précédent livre et celui-ci. Cette remarque me permet d’insister tout d’abord sur le fait que ce livre n’est pas une paraphrase du précédent (heureusement !), et ensuite sur le fait que la science avance, et que ces modèles successifs illustrent comment on passe d’une situation simple à une situation plus compliquée, et le temps que cela prend. A la date d’aujourd’hui, la question qui m’intéresse est de savoir comment les capillaires se décrochent des vaisseaux moyens ou gros, et si la mécanique du tissu sous pression entre les tuyaux, ainsi que la présence de fibres (encore elles) n’y jouent pas un rôle essentiel. Pour finir, regardons une dernière fois une noix, un cerveau, les ruisseaux de la plage du Plat Gousset, la peau du shar-peï et les veines de nos vingt ans : toutes ces formes ont en commun d’être désordonnées. Bien qu’il y ait autant sinon davantage de formes désordonnées qu’ordonnées dans la nature, notre œil est particulièrement sensible aux formes régulières. Mais le chien est plié pratiquement au hasard, les arborescences de Granville branchent de même et tout cela n’est pas très géométriquement ordonnancé, cher monsieur Galilée.