La spéciation géographique : La spéciation géographique par fractionnement de l'aire de répartition
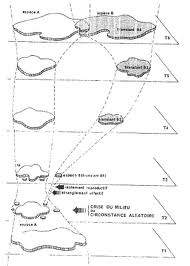 Le modèle mésange permet d’illustrer deux points. Tout d’abord, une espèce est constituée par un ensemble de populations locales plus ou moins génétiquement isolées les unes des autres. Les individus qui composent ces populations sont capables d’accouplement et ils occupent un espace délimité. Ces individus partagent un même pool de gènes et dans l’idéal c’est une unité panmictique, c’est-à-dire que les individus qui la composent s’accouplent au hasard. Quand la divergence entre deux ensembles de populations de la même espèce atteint un certain niveau, on qualifie ces deux ensembles de sous-espèces. Ce niveau de différenciation correspond en général à un isolement reproducteur mais les individus de chaque sous- espèce peuvent encore sans ambiguïté être classés dans la même espèce. C’est le cas des mésanges bleues d’Afrique du Nord par rapport à celles d’Europe occidentale continentale. Ces différentes structurations de l’espèce sont données . Ensuite, deux types de spéciations semblent possibles au terme de cette étude : la spéciation par fractionnement d’aire de répartition et la spéciation par colonisation d’île. Peut-on établir ces types de spéciation avec précision ? Ces deux spéciations résultent de l’isolement d’une fraction plus ou moins importante de la population par la mise en place d’une barrière géographique. Ce sont donc des spéciations allopatriques. Des spéciations sympatriques sont-elles possibles ?Chez les oiseaux on connaît différents exemples d’espèces ou semi-espèces dont l’apparition peut être expliquée par le fractionnement de l’aire de répartition d’une espèce originelle. Le plus connu est sans doute celui des Corneilles noire et mantelée Corvus corone corone et Corvus corone cornix (voir Mayr 1974) qui se partagent l’Europe selon une ligne à peu près nord-sud . Dans la zone de contact entre ces deux semi-espèces des couples mixtes féconds existent mais leur existence semble limitée à cette zone de contact au demeurant peu profonde. On peut proposer le modèle suivant pour l’apparition de ces deux semi-espèces. Lors des glaciations quaternaires, l’aire de répartition d’une espèce de corneille habitant ‘ Europe a été séparée en deux par la progression des glaciers vers le sud. On peut situer les deux zones où se sont réfugiées les corneilles dans la péninsule ibérique et dans les Balkans.En situation allopatrique, ces deux populations ont pu diverger l’une par rapport à l’autre. Le retrait des glaces a permis à ces populations de recoloniser le terrain perdu. Cependant la divergence entre ces deux groupes n’ayant pas conduit à un changement de niche, ils ne cohabitent pas. Le processus de spéciation s’est donc arrêté en chemin. Le problème posé est bien entendu de déterminer le(s) facteur(s) empêchant ces deux semi-espèces de cohabiter. On admet généralement que c’est la compétition due à leur utilisation des même ressources du milieu. Nous discuterons plus loin de la validité de cet argument. Cependant, cet exemple ô combien pédagogique, s’avère plus complexe qu’il n’y paraît lorsque l’on s’intéresse à l’ensemble de l’aire de répartition de la Corneille noire. En effet, celle-ci est également présente en Extrême-Orient ce qui ne facilite pas l’interprétation, à moins de supposer que quelques individus ont accompagné Marco Polo lors de son voyage en Chine et ont fondé la population orientale ! ! ! Plus sérieusement, une étude approfondie de la divergence génétique entre les individus de toutes les populations permettrait peut-être de montrer que les individus d’Extrême-Orient appartiennent à une autre sous-espèce. Il faudrait alors envisager non pas deux mais trois zones refuge ayant fonctionné simultanément pendant les périodes glaciaires.Les perdrix méditerranéennes du genre Alectoris présentent un autre exemple de spéciation par fractionnement (détails dans Blondel 1995). On connaît dans le bassin méditerranéen 4 espèces de perdrix appartenant à ce genre : la Perdrix rouge (Alectoris rufa), la Perdrix bartavelle (Alectoris graeca), la Perdrix chukar (Alectoris chukar) et la Perdrix gambra (Alectoris barbara). Ces quatre espèces sont des oiseaux des biotopes ouverts, secs et plus ou moins rocailleux.D’un point de vue paléontologique (Mourer-Chauviré 1975), on connaît au Pléistocène des fossiles proches de la Perdrix bartavelle actuelle. C’est à l’Holocène qu’apparaissent des fossiles de Perdrix rouge. Par contre, on connaît des fossiles de Perdrix gambra dès le Pléistocène moyen simultanément à des fossiles de Perdrix bartavelle.
Le modèle mésange permet d’illustrer deux points. Tout d’abord, une espèce est constituée par un ensemble de populations locales plus ou moins génétiquement isolées les unes des autres. Les individus qui composent ces populations sont capables d’accouplement et ils occupent un espace délimité. Ces individus partagent un même pool de gènes et dans l’idéal c’est une unité panmictique, c’est-à-dire que les individus qui la composent s’accouplent au hasard. Quand la divergence entre deux ensembles de populations de la même espèce atteint un certain niveau, on qualifie ces deux ensembles de sous-espèces. Ce niveau de différenciation correspond en général à un isolement reproducteur mais les individus de chaque sous- espèce peuvent encore sans ambiguïté être classés dans la même espèce. C’est le cas des mésanges bleues d’Afrique du Nord par rapport à celles d’Europe occidentale continentale. Ces différentes structurations de l’espèce sont données . Ensuite, deux types de spéciations semblent possibles au terme de cette étude : la spéciation par fractionnement d’aire de répartition et la spéciation par colonisation d’île. Peut-on établir ces types de spéciation avec précision ? Ces deux spéciations résultent de l’isolement d’une fraction plus ou moins importante de la population par la mise en place d’une barrière géographique. Ce sont donc des spéciations allopatriques. Des spéciations sympatriques sont-elles possibles ?Chez les oiseaux on connaît différents exemples d’espèces ou semi-espèces dont l’apparition peut être expliquée par le fractionnement de l’aire de répartition d’une espèce originelle. Le plus connu est sans doute celui des Corneilles noire et mantelée Corvus corone corone et Corvus corone cornix (voir Mayr 1974) qui se partagent l’Europe selon une ligne à peu près nord-sud . Dans la zone de contact entre ces deux semi-espèces des couples mixtes féconds existent mais leur existence semble limitée à cette zone de contact au demeurant peu profonde. On peut proposer le modèle suivant pour l’apparition de ces deux semi-espèces. Lors des glaciations quaternaires, l’aire de répartition d’une espèce de corneille habitant ‘ Europe a été séparée en deux par la progression des glaciers vers le sud. On peut situer les deux zones où se sont réfugiées les corneilles dans la péninsule ibérique et dans les Balkans.En situation allopatrique, ces deux populations ont pu diverger l’une par rapport à l’autre. Le retrait des glaces a permis à ces populations de recoloniser le terrain perdu. Cependant la divergence entre ces deux groupes n’ayant pas conduit à un changement de niche, ils ne cohabitent pas. Le processus de spéciation s’est donc arrêté en chemin. Le problème posé est bien entendu de déterminer le(s) facteur(s) empêchant ces deux semi-espèces de cohabiter. On admet généralement que c’est la compétition due à leur utilisation des même ressources du milieu. Nous discuterons plus loin de la validité de cet argument. Cependant, cet exemple ô combien pédagogique, s’avère plus complexe qu’il n’y paraît lorsque l’on s’intéresse à l’ensemble de l’aire de répartition de la Corneille noire. En effet, celle-ci est également présente en Extrême-Orient ce qui ne facilite pas l’interprétation, à moins de supposer que quelques individus ont accompagné Marco Polo lors de son voyage en Chine et ont fondé la population orientale ! ! ! Plus sérieusement, une étude approfondie de la divergence génétique entre les individus de toutes les populations permettrait peut-être de montrer que les individus d’Extrême-Orient appartiennent à une autre sous-espèce. Il faudrait alors envisager non pas deux mais trois zones refuge ayant fonctionné simultanément pendant les périodes glaciaires.Les perdrix méditerranéennes du genre Alectoris présentent un autre exemple de spéciation par fractionnement (détails dans Blondel 1995). On connaît dans le bassin méditerranéen 4 espèces de perdrix appartenant à ce genre : la Perdrix rouge (Alectoris rufa), la Perdrix bartavelle (Alectoris graeca), la Perdrix chukar (Alectoris chukar) et la Perdrix gambra (Alectoris barbara). Ces quatre espèces sont des oiseaux des biotopes ouverts, secs et plus ou moins rocailleux.D’un point de vue paléontologique (Mourer-Chauviré 1975), on connaît au Pléistocène des fossiles proches de la Perdrix bartavelle actuelle. C’est à l’Holocène qu’apparaissent des fossiles de Perdrix rouge. Par contre, on connaît des fossiles de Perdrix gambra dès le Pléistocène moyen simultanément à des fossiles de Perdrix bartavelle.
Tout ceci permet de supposer qu’un taxon ancien (bartavelle) a vu son aire de répartition fractionnée par les événements climatiques du Quaternaire. Les travaux des paléobotanistes montrent que des îlots de végétation méditerranéenne ont existé au cœur des glaciations sur les berges de la Méditerranée . Les populations temporairement isolées ont génétiquement divergé puis se sont trouvées en contact secondaire lors du retrait des glaciers. Le processus de différenciation n’a cependant pas été suffisant et l’isolement reproducteur n’existe pas. De même les niches écologiques de ces différentes perdrix son restées quasi identiques. Il s’agit de semi-espèces et les Perdrix rouge, bartavelle et chukar constituent une super-espèce.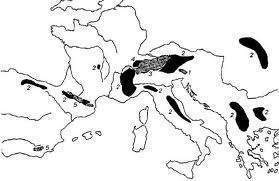 On notera cependant que les niches écologiques de la Perdrix rouge et de la Perdrix bartavelle diffèrent dans les Alpes Maritimes françaises, là où elles sont en contact. En effet, les bartavelles sont là des oiseaux d’altitude atteignant les pelouses de l’étage alpin, alors que les Perdrix rouges se cantonnent aux zones de basse altitude. Par contre, en Grèce, là où la Perdrix rouge est absente, la Perdrix bartavelle vit à basse altitude, dans l’habitat occupé par la Perdrix rouge dans les Alpes Maritimes. On peut faire appel à la compétition pour expliquer ce fait. Nous en reparlerons.
On notera cependant que les niches écologiques de la Perdrix rouge et de la Perdrix bartavelle diffèrent dans les Alpes Maritimes françaises, là où elles sont en contact. En effet, les bartavelles sont là des oiseaux d’altitude atteignant les pelouses de l’étage alpin, alors que les Perdrix rouges se cantonnent aux zones de basse altitude. Par contre, en Grèce, là où la Perdrix rouge est absente, la Perdrix bartavelle vit à basse altitude, dans l’habitat occupé par la Perdrix rouge dans les Alpes Maritimes. On peut faire appel à la compétition pour expliquer ce fait. Nous en reparlerons.
Pour la Perdrix gambra la présence de fossiles de formes voisines au Pléistocène permet de dire qu’il s’agit d’un groupe ancien mais l’absence de zones de contact avec les autres perdrix ne permet pas de statuer sur le niveau d’isolement reproductif. On peut seulement dire que la séparation avec les autres perdrix est plus ancienne.
On trouvera dans la littérature d’autres exemples de fractionnements d’aires de répartition sous l’effet de changements climatiques et conduisant à des semi-spéciations ou à des spéciations. Blondel (1995) et Ridley (1997) présentent un tel modèle pour différentes espèces de la forêt amazonienne (travaux de Haffer et Simpson et travaux de Haffer) où existent de nombreux foyers d’endémisme attribués à un fractionnement de la forêt à l’époque glaciaire. Dans chaque îlot forestier, les populations ainsi temporairement isolées ont eu le temps de diverger génétiquement les unes par rapport aux autres. On trouvera d’autres exemples dans Blondel (1995) concernant des oiseaux de la Cordillère des Andes et des traquets eurasiatiques et dans Mayr (1974) pour des sittelles d’Australie. L’importance des glaciations quaternaires dans les phénomènes de spéciation chez les oiseaux a cependant été discutée. Ainsi, des études palynologiques (Colinvaux 1979) montrent qu’au moins une zone considérée par Haffer comme sèche durant la dernière glaciation dans le bassin amazonien est restée au con¬traire forestière durant toute cette période. De plus, Haffer a reconstitué les zones refuges en utilisant les richesses en espèces d’oiseaux ainsi que les données pluviométriques actuelles. De même une étude de l’ADN mitochondrial de 35 passereaux d’Amérique du Nord (Klicka & Zink 1997) montre que ces espèces divergent 10 fois plus que ce que laisserait prévoir une spéciation survenue à la suite des glaciations du Pléistocène. Ces distances génétiques doivent cependant être prises avec précaution. En effet, des travaux récents (Strauss 1999) montrent que des recombinaisons entre ADN mitochondrial d’origine mâle et d’origine femelle peuvent se produire après la fécondation. Ces événements, en introduisant une source de divergence non prise en compte dans les calculs, font que la distance génétique réelle est peut-être supérieure à celle qui était calculée jusqu’alors. Les divergences entre les ADN mitochondriaux des passereaux étudiés ne sont donc peut-être pas incompatibles avec une spéciation récente.