La régionalisation du corps
La régionalisation du corps
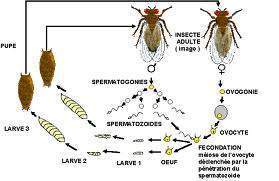
Pour bien fixer les idées nous allons faire un petit tour chez la drosophile où les modèles ont été préalablement établis puis transférés aux Vertébrés (Alberts et al 1994, Gilbert 1996).
Les premiers gènes intervenant dans le développement embryonnaire sont des gènes maternels. Ils déterminent la polarité de l’œuf grâce à des produits fournis par les cellules nourricières du follicule et localisés dans le cytoplasme de l’ovocyte. C’est là une occasion supplémentaire de rappeler l’importance de l’hérédité cytoplasmique. La polarité dorso- ventrale est le résultat de l’action d’une vingtaine de produits maternels, agissant en cascade aboutissant à la formation d’un facteur de transcription, la protéine dorsal. Celle-ci est distribuée dans tout le cytoplasme du zygote mais ne pénétrera que dans les noyaux des cellules ventrales de l’embryon, y activant les gènes ventralisant (,snail et twist) et y inhibant les gènes dorsalisant (decapentaplegic et zerhüilt).
Préalablement à cela, la polarité antéro-postérieure est mise en place grâce à des ARN distribués de façon plus ou moins homogène dans le cytoplasme de l’ovocyte. Cela concerne les ARN des gènes bicoïd, hunchback et nanos, encore qualifiés de gènes à effet maternel. L’ARNm bicoïd est présent dans le cytoplasme antérieur, l’ARNm nanos dans le cytoplasme postérieur et l’ARNm hunchback est présent partout. Après la fécondation, les ARNm bicoïd et nanos sont traduits en protéines qui sont des facteurs de régulation de l’expression du gène hunchback. La protéine bicoïd active la transcription du gène hunchback de l’embryon alors que la protéine nanos inhibe la traduction de l’ARNm hunchback. Cette protéine est elle même un répresseur de l’expression des gènes abdominaux ; ceux-ci peuvent donc s’exprimer uniquement dans la région postérieure de l’embryon. D’élégantes manipulations démontrent cette fonction régionalisante : le remplacement du cytoplasme antérieur d’un zygote par le cytoplasme postérieur d’un ovocyte conduit au développement d’un embryon présentant une région postérieure à chaque extrémité.
La segmentation de l’embryon est ensuite contrôlée par des groupes de gènes de segmentation dont l’expression dépend des gradients précédents. Il s’agit des groupes gap (hunchback, Kriippel, knirps, giant, …), pairrule (hairy, even-skipped, paired, fushi- tarazu…) et des gènes de polarité de segment (engrailed, wingless, hedgehog, …). L’expression des gènes gap est activée ou réprimée par les produits des gènes à effet maternel et délimitent les grandes régions de l’embryon. Les gènes pair-rule sont activés ou réprimés par les produits des gènes gap et découpent les régions précédentes en segments. Enfin, les gènes de polarité de segment régionalisent chacun des segments formés précédemment, c’est- à-dire qu’ils y déterminent une région antérieure et une région postérieure. L’embryon est alors « découpé » en un nombre fixe de segments et de parasegments (2 demi-segments contigus). Le passage d’un gradient de substances à une distribution en bandes est réalisé par la fixation coopérative des facteurs de transcription sur les différents promoteurs des gènes cibles.
Ce découpage permet ensuite une activation séquentielle des gènes homéotiques, regroupés dans les ensembles Antennapedia et Bithorax du complexe HOM situé sur le chromosome 3 . Ces gènes sont activés ou réprimés par les produits des gènes pair-rule et gap et inhibés par les produits des gènes homéotiques qui leur sont postérieurs. Il existe donc une autorégulation du système. Chaque segment fabrique alors, entre autre, une paire d’appendice en fonction de sa localisation.
Les gènes homéotiques codent pour des protéines activatrices du génome, des facteurs de transcription, qui interagissent avec l’ADN par un homéodomaine de 60 acides aminés contenant 3 segments d’hélice alpha associés en un motif hélice-tour-hélice. Cet homéodomaine est codé par une séquence de 180 paires de bases appelée homéoboîte. l’interaction concerne essentiellement des séquences activatrices de gènes appelés « gènes réalisateurs », responsables de la détermination tissulaire. Ces gènes effecteurs sont activement étudiés à l’heure actuelle et restent pour l’instant mal connus, mais peuvent être des gènes de structure aussi bien que d’autres gènes homéotiques. Il y a donc une organisation hiérarchique dans l’expression et la régulation du génome, les gènes « homéotiques » étant des commutateurs/régulateurs du développement. On se trouve en présence d’une interaction de cascades régulatrices subissant chacune une autorégulation positive ou négative.
La situation semble très comparable chez les Vertébrés tout au moins en ce qui concerne l’édification des axes, grâce entre autre au gène goosecoïd, et les gènes homéotiques . Le plus surprenant est que des gènes homéotiques ont été retrouvés contrôlant ¦e développement d’organismes aussi différents que les Insectes, les Amphibiens ou les Mammifères, ou encore d’animaux à corps non segmenté, Nématodes, par exemple, et même de plantes, lors de la mise à fleurs c’est-à-dire au moment de l’établissement de l’identité des différentes pièces florales.
Cette organisation renforce si besoin était l’idée d’une origine commune à tous les êtres vivants, et amène à retracer quelques étapes de l’histoire. En effet, la comparaison des séquences d’acides aminés des protéines codées par ces gènes permet d’estimer le degré de parenté qui les relie . Dans le cas des gènes homéotiques, la séquence subissant la plus forte contrainte fonctionnelle, l’homéoboîte ou plus exactement sa version traduite, l’homéodomaine, reste très conservée alors que les séquences voisines sont très variables. C’est la raison pour laquelle cette séquence particulière fait l’objet de ces études comparatives. La figure 4.20 comporte les structures primaires des homéodomaines de 6 des protéines codées par le complexe HOM de la drosophile et de 6 des protéines occupant les mêmes positions relatives au sein du complexe HoxB de la souris. Il est intéressant de noter que ces deux complexes, bien qu’appartenant à des espèces très éloignées au plan phylétique, possèdent le même nombre de gènes. Au sein d’un même complexe le degré d’homologie entre séquences conduit à penser à une série de duplications géniques successives comme mécanisme de formation de ces complexes. Mais, entre espèces, on constate que, par exemple, les séquences des homéodomaines des protéines Antp, de la drosophile, et HoxB-6, de la souris, diffèrent de seulement 4 acides aminés. C’est-à-dire qu’il y a plus d’homologie entre deux séquences occupant la même place au sein des complexes de deux espèces très différentes qu’entre deux séquences occupant des places différentes au sein du même complexe. On a donc l’impression que les complexes se sont mis en place il y a très longtemps, avant la divergence des groupes que l’on étudie aujourd’hui. La variation du nombre de gènes au sein de chaque complexe est en revanche due à des duplications ultérieures, de même que l’augmentation du nombre de complexes dans certains groupes. Chaque complexe évoluant ensuite pour son propre compte.Une telle organisation hiérarchique de la régulation de l’expression du génome permet de percevoir l’effet de mutations ponctuelles, ou d’inactivations de ces gènes. La structure du plan d’organisation pourrait en être profondément modifiée. On pense en particulier (Gilbert 1996) que la mise en place d’une métamérie hétéronome et la tagmatisation, c’est-à-dire l’association de métamères contigus au sein d’unités structurales et fonctionnelles (tête, thorax, abdomen), chez les ancêtres de la drosophile, sont dus à des duplications et divergences successives des gènes homéotiques. De la même façon, on a montré que des mutations de ces mêmes gènes pouvaient faire resurgir des structures ancestrales. C’est le cas pour le gène HoxA-2 de la souris pour lequel une délétion peut faire réapparaître des structures reptiliennes de la mâchoire inférieure (Rijli et al. 1993).
Vidéo: La régionalisation du corps
Vidéo démonstrative pour tout savoir sur: La régionalisation du corps