Vers une définition opérattionnelle de l'espèce
 La notion d’espèce est ancienne et remonte à l’aube de l’humanité. Les premiers chasseurs cueilleurs ont ainsi nommé les animaux et les végétaux dont ils se nourrissaient. On constate d’ailleurs que la correspondance est assez bonne, tout au moins pour celles de grande taille et en particulier celles qui présentent un intérêt alimentaire, entre les espèces reconnues par les populations humaines de diverses régions du monde et les espèces identifiées par les équipes de scientifiques occidentaux qui ont visité ces mêmes régions. Ainsi, Mayr rapporte que les Papous de Nouvelle-Guinée ont 136 noms pour les 137 espèces d’oiseaux qu’il avait identifiées. Les Kalams de Nouvelle-Guinée reconnaissent 174 espèces de vertébrés là où les scientifiques occidentaux en identifient 170. Les Fores, autre peuplade de Nouvelle-Guinée, mis en présence d’espèces d’oiseaux qu’ils n’avaient jamais rencontrés, les placent dans presque 90% des cas et sans hésitation dans le même groupe défini par les occidentaux que des espèces qu’ils connaissent (voir Gould 1982, auquel sont empruntés ces exemples, pour d’autres cas de telles correspondances). Comme le fait remarquer Mayr, si des peuplades dites primitives reconnaissent les mêmes subdivisions de la nature que des scientifiques formés à l’université, alors l’espèce est bien une subdivision naturelle du vivant et c’est la seule qui a une signification biologique. Comment la définir ?
La notion d’espèce est ancienne et remonte à l’aube de l’humanité. Les premiers chasseurs cueilleurs ont ainsi nommé les animaux et les végétaux dont ils se nourrissaient. On constate d’ailleurs que la correspondance est assez bonne, tout au moins pour celles de grande taille et en particulier celles qui présentent un intérêt alimentaire, entre les espèces reconnues par les populations humaines de diverses régions du monde et les espèces identifiées par les équipes de scientifiques occidentaux qui ont visité ces mêmes régions. Ainsi, Mayr rapporte que les Papous de Nouvelle-Guinée ont 136 noms pour les 137 espèces d’oiseaux qu’il avait identifiées. Les Kalams de Nouvelle-Guinée reconnaissent 174 espèces de vertébrés là où les scientifiques occidentaux en identifient 170. Les Fores, autre peuplade de Nouvelle-Guinée, mis en présence d’espèces d’oiseaux qu’ils n’avaient jamais rencontrés, les placent dans presque 90% des cas et sans hésitation dans le même groupe défini par les occidentaux que des espèces qu’ils connaissent (voir Gould 1982, auquel sont empruntés ces exemples, pour d’autres cas de telles correspondances). Comme le fait remarquer Mayr, si des peuplades dites primitives reconnaissent les mêmes subdivisions de la nature que des scientifiques formés à l’université, alors l’espèce est bien une subdivision naturelle du vivant et c’est la seule qui a une signification biologique. Comment la définir ?
Chez les scientifiques, le concept typologique de l’espèce a longtemps prévalu. C’est celui de Platon, Aristote, Linné, pour lesquels on définit l’espèce par un type, c’est-à-dire un représentant conservé en musée et soigneusement décrit. Un individu ne peut s’écarter du type que par accident ou suite à une observation défectueuse. Cette définition est fixiste et elle ignore le polymorphisme. Les premiers scientifiques à avoir étudié l’espèce se limitaient ainsi à dresser des inventaires. C’était la grande époque où les naturalistes européens, puis nord américains, ramenaient de leurs voyages aux quatre coins de la planète des exemplaires d’espèces animales et végétales nouvelles pour la science occidentale. Ces spécimens étaient décrits et classés, donnant naissance aux fameuses collections de nos muséums. Entre le XVIIe et le XIXe siècle, cet acharnement à décrire répondait à deux aspirations. Les scientifiques étaient émerveillés par la diversité et l’harmonie du vivant et nombre d’entre eux ne con¬cevaient pas qu’une telle harmonie puisse être autre chose que l’expression directe de la volonté divine. La classification et l’étude de la diversité du vivant représentaient pour eux des moyens d’accéder à la connaissance de Dieu. D’autres scientifiques pensaient au contraire établir par la classification des lois universelles du type de celles que l’on découvrait alors en physique par exemple et qui auraient permis d’expliquer cette diversité du vivant (voir Mayr 1989 pour l’historique de la notion d’espèce).
Cette approche typologique de l’espèce n’est cependant pas satisfaisante. D’une part les individus d’une même espèce présentent des différences morphologiques sur l’étendue de leur aire de répartition et on distingue ainsi des populations géographiques comme l’étude de la Mésange bleue nous l’a montré. Ensuite, au sein d’une population, les individus présentent des différences morphologiques : il y a polymorphisme. Si le dimorphisme sexuel ou l’existence de stades larvaires sont des problèmes assez facilement surmontés, encore que Linné avait fait des canards colvert mâle et femelle deux espèces distinctes, les espèces jumelles, quasi indiscernables morphologiquement, marquent bien la limite du concept typologique dans la définition de l’espèce. C’est le cas des souris Mus spretus et Mus musculus qui ne diffèrent que par la longueur de leur queue, des Mésanges nonnette (Parus palustris) et boréale {Parus montanus) ou des Hypolaïs polyglotte (Hippolais polyglotta) et ictérine {Hippolais icterina) qui sont des espèces d’oiseaux que l’on ne différencie bien que par le chant.
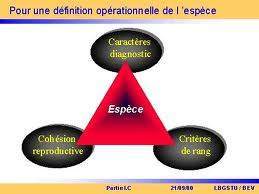
L’introduction de la notion de reproduction permet de résoudre partiellement ce problème. Deux individus appartiennent à la même espèce s’ils se reproduisent entre eux mais pas avec des individus d’autres espèces. Ce critère est assez ancien puisqu’on le trouve chez John Ray qui écrivait en 1680 : « Une espèce ne sort jamais de la semence d’une autre espèce
et vice-versa » et chez Cuvier qui définissait l’espèce en 1812 comme « une collection de tous les corps nés les uns des autres ou de parents communs et qui leur ressemblent autant qu’ils se ressemblent entre eux ». Dans l’esprit de certains biologistes la reproduction sexuée permettait la transmission au cours du temps de l’essence de l’espèce qui venait de la création. Le concept d’espèce est donc indissociable de celui de reproduction sexuée. Le brassage génétique qu’elle induit est responsable de la ressemblance entre les individus appartenant à une même espèce. Quant à une population qui ne recourt qu’à la reproduction asexuée, elle ne constituerait pas une espèce mais autant d’espèces qu’elle comporte de clones génétiquement distincts. La définition la plus souvent citée de l’espèce intégrant le critère d’interfécondité émane cependant de l’éminent zoologiste Emst Mayr qui l’a définie en 1942 comme «un groupe de populations naturelles actuellement ou potentiellement interfécondes et reproductive-ment isolées des autres groupes équivalents ». C’est le concept biologique de l’espèce.
Certaines critiques amenèrent Mayr à supprimer le concept de potentiellement interfécondes dans sa définition en 1963 (1974 pour l’édition française) et d’y ajouter la dimension écologique en 1982 (1989 pour l’édition française). Ainsi, dorénavant, «une espèce est une communauté reproductive de populations, reproductivement isolée d’autres communautés, qui occupe une niche particulière dans la nature ». Cet ajout à la définition de 1942 est loin d’être anodin car il donne tout son sens à l’isolement reproducteur. De nombreuses études suggèrent que dans une aire géographique donnée, le nombre de niches est à peu près fixé et une niche n’accueille donc qu’une espèce (voir chapitre 3.1. pour la définition de la niche écologique). Par contre, la nature des espèces qui occupent chaque niche reflète le hasard des processus évolutifs et de colonisation. On peut avancer comme argument le fait que l’introduction d’une espèce étrangère en élimine une au moins parmi celles déjà en place et en priorité celle qui occupe une niche voisine de celle de l’envahisseur. Le concept biologique est une définition génétique de l’espèce qui peut donc être caractérisée par certaines associations de gènes (c’est un pool de gènes). L’isolement reproducteur permet d’éviter que cette association harmonieuse de gènes, indispensable à l’exploitation de la niche, et donc à la survie des individus et de leurs descendants, ne soit perturbée par des gènes issus d’autres espèces, dont le pool de gènes est adapté à d’autres conditions écologiques.
On notera que, curieusement, les ouvrages français sur l’évolution et tous les manuels scolaires reprennent la définition biologique de l’espèce de Mayr mais ils se limitent à celles de 1942 ou de 1963 et aucun n’introduit la dimension écologique. Même les ouvrages qui démontrent que la spéciation correspond à la conquête de nouvelles niches écologiques se .mitent au critère d’interfécondité dans leur définition de l’espèce. On est tenté de rapprocher ce constat du jugement que le généticien Richard Lewontin avait porté sur la France dans une interview accordée au journal Le Monde en 1980 et dans laquelle il déclarait : «je n’irai pas jusqu’à dire que, dans ce pays, la pensée évolutionniste a des années de retard, mais enfin, il y a beaucoup à faire». Gouyon, Henry & Amould (1997) font un constat similaire dans l’introduction de leur ouvrage lorsqu’ils stigmatisent la résistance au darwinisme de la communauté scientifique de l’hexagone.
Cette dernière définition de Mayr qui a notre préférence a aussi été critiquée, en particulier à cause de la difficulté à définir la niche et la communauté reproductive. On notera que ceux qui la critiquent ne sont souvent pas des biologistes des populations. Ils sont de ce fait peu familiers avec le concept de niche qui doit être conçu comme un espace multidimensionnel. Mais cela ne signifie pas que définir une espèce est aisé. La naissance de nouvelles espèces, processus bien documenté de nos jours quand il résulte d’un isolement reproducteur préalable comme nous l’avons montré dans les paragraphes qui précèdent, est généralement considérée comme lente. Lors de ce processus, des populations de la même espèce deviennent peu à peu au cours du temps des sous-espèces puis des espèces différentes. Suivant à quel stade on étudie le processus, on ne peut pas établir l’existence de deux espèces distinctes. Définir à tout prix des espèces revient parfois à vouloir fixer une limite là où n’existe qu’un continuum. Il en est de même lorsque l’on cherche à définir une espèce en paléontologie. En
dehors du manque de critères utilisables lié à un matériel généralement incomplet, on peut se demander à quel moment on passe d’une espèce à une autre au cours d’un processus évolutif. Certains paléontologues ont ainsi défini l’espèce dans le temps en appliquant ce terme à des lignées évolutives indépendantes d’autres lignées et inféodées à une zone adaptative donnée. Mais ces problèmes n’ont guère d’importance, car la vraie question n’est pas de définir des espèces (l’intérêt en est limité). Le vrai problème est de comprendre les mécanismes de la spéciation, que l’on peut définir comme l’acquisition de l’isolement reproducteur permettant la conquête d’une nouvelle niche écologique et qui constitue le premier palier de l’évolution. L’acquisition de l’isolement reproducteur est donc une étape fondamentale dans le processus de spéciaticTl. Les barrières qui maintiennent cet isolement sont de diverses natures et elles peuvent être regroupées en deux ensembles. Les barrières prézygotiques empêchent la fécondation et donc la formation d’un zygote. Elles peuvent être écologiques (les espèces vivent dans des habitats différents), phénologiques (les périodes de reproduction des espèces ne sont pas synchrones) comportementales (parades nuptiales, phéromones ou chants différents), anatomiques (incompatibilité des organes génitaux) ou gamétiques (arrêt de croissance des tubes polliniques, mort des spermatozoïdes dans les voies génitales de la femelle, non reconnaissance entre les gamètes). Les barrières postzygotiques interviennent après la fécondation. Elles concernent les développements embryonnaires qui avortent et les cas où les hybrides sont stériles ou ont une fécondité ou une viabilité réduite.