L'influence de la prédation sur la dynamique des populations : Les limites de l'influence de la prédation
 Comme nous l’avons montré pour la compétition, la prédation est parfois invoquée à tort comme facteur causal de processus évolutifs. Ceci est tout à fait clair sur certains modèles trop pédagogiques pour être tout à fait exacts. C’est le cas des variations d’effectif dans les systèmes prédateur-proie. L’examen des courbes de variation des effectifs de « proies » et de «prédateurs » amène quelques remarques : on voit ici nettement que ces variations sont loin d’être régulières et auraient plutôt un aspect « chaotique », c’est-à-dire vraisemblablement gouvernées par des processus déterministes, mais dont la sensibilité aux conditions initiales du système rend toute prédiction à long terme quasi impossible (May 1991, Ruelle 1991).
Comme nous l’avons montré pour la compétition, la prédation est parfois invoquée à tort comme facteur causal de processus évolutifs. Ceci est tout à fait clair sur certains modèles trop pédagogiques pour être tout à fait exacts. C’est le cas des variations d’effectif dans les systèmes prédateur-proie. L’examen des courbes de variation des effectifs de « proies » et de «prédateurs » amène quelques remarques : on voit ici nettement que ces variations sont loin d’être régulières et auraient plutôt un aspect « chaotique », c’est-à-dire vraisemblablement gouvernées par des processus déterministes, mais dont la sensibilité aux conditions initiales du système rend toute prédiction à long terme quasi impossible (May 1991, Ruelle 1991).
On peut, à cette occasion, nuancer l’interprétation que l’on donne d’un exemple classique concernant les variations d’effectifs de populations sympatriques de Lynx et de Lièvre américain à Terre-Neuve. Cet exemple résulte de la combinaison de deux sources : un article de Bergerud (1984) et un travail plus ancien. L’explication simple (simpliste?) qui vient immédiatement à l’esprit lorsque cette courbe est fournie sans commentaire, est que plus il y a de lièvres, plus il y a de lynx (abondance de proies) qui font alors chuter les effectifs de lièvres (prédation forte) et dont les effectifs diminuent en conséquence (carence de proies) ce qui a pour effet d’alléger la prédation subie par la population de lièvres qui se met à proliférer, et c’est reparti pour un tour.
Le « hic », c’est que ces données, en particulier les plus anciennes, sont obtenues à partir de l’analyse de comptes rendus de chasse. Or les chasseurs, comme les lynx, consomment du lièvre pendant leurs campagnes. On peut donc « raconter » une autre histoire : plus il y a de lièvres, plus les chasseurs peuvent faire des réserves alimentaires et intensifier leur effort de chasse au lynx. Le nombre de peaux rapportées augmente donc sans que l’on sache précisément comment ont varié les effectifs de cette espèce. Les « variations » des deux populations naturelles ne sont peut-être pas aussi strictement corrélées qu’on veut bien le dire !
Nous n’affirmons pas pour autant qu’il n’y a pas de corrélation entre effectifs de populations de proies et de prédateurs. En effet, des travaux récents effectués dans des conditions expérimentales très strictes, montrent pour cet exemple que l’élimination des prédateurs est suivie d’un doublement de la densité des lièvres mais que dans le même temps l’apport de nourriture supplémentaire, sans variation de la densité des prédateurs, triple la densité des lièvres. Lorsque l’on combine suppression des prédateurs et apport de nourriture, la densité des proies est multipliée par 11 dans les parcelles expérimentales (Krebs et al. 1995). Les effectifs des populations apparaissent donc bien liés à plusieurs facteurs simultanément. D’autre part, ces expérimentations ne disent pas si la prédation est la cause des cycles ou si d’autres facteurs en sont éventuellement responsables, le parasitisme par exemple. Cette situation est en effet documentée chez des chenilles et chez des rongeurs et la prudence s’impose donc avant toute interprétation catégorique.
On peut cependant faire apparaître des variations d’effectif finement corrélées dans un système prédateur-proie très simplifié. C’est le cas lorsqu’on élève ensemble des Protozoaires dans des boîtes de Pétri. Encore faut-il respecter des conditions très strictes (limitation des contacts, etc.) pour assurer la coexistence des deux espèces pendant un temps supérieur à quelques jours (Barbault 1981). Le cas des populations naturelles est beaucoup plus complexe. Cependant, dans tous les cas et quels que soient les mécanismes, les effectifs des populations de proies et de prédateurs passent par des valeurs minimales, ce qui diminue le nombre de reproducteurs et donc appauvrit le pool génique de la population et sa variabilité. Des phénomènes de dérive génétique peuvent donc se mettre en place et provoquer ou accentuer des processus évolutifs.
Compte tenu de ces remarques, on peut conclure que l’impact d’un prédateur ou d’un parasitoïde sur une population de proies ou d’hôtes n’est qu’un des éléments du complexe sélectif global constitué de l’ensemble des interactions permettant de définir un système population-environnement. Ce système prend en compte l’ensemble des relations établies entre la population et son environnement biotique et abiotique. 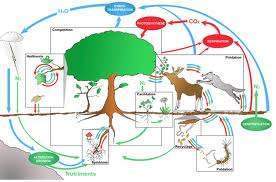 L’influence de la prédation sur l’évolution d’une population de proies sera donc plus ou moins important dans la mesure où la structure génétique de cette population représente un compromis adaptatif.
L’influence de la prédation sur l’évolution d’une population de proies sera donc plus ou moins important dans la mesure où la structure génétique de cette population représente un compromis adaptatif.