L'organisation spatiale de l'espèce : Approche biométrique
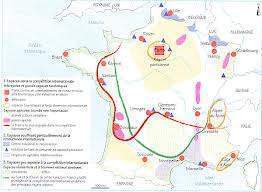
 L’étude de la variation géographique a donc été pour Darwin un élément important de réflexion dans la mise au point de sa théorie. Nous allons tout d’abord montrer comment de telles études de populations permettent de proposer des modèles de spéciation acceptables. Pour cela, nous allons exploiter un ensemble de travaux menés au Centre L. Emberger (C.N.R.S., Montpellier) par l’équipe de J. Blondel sur la Mésange bleue {Parus caeruleus). L’aire de répartition de cette espèce comprend les îles Canaries, l’Afrique du Nord, la Cyrénaïque, toute l’Europe jusqu’au sud de la Scandinavie au nord et jusqu’à l’Oural à l’est, la Turquie et une partie de l’Iran. Sur l’ensemble de son aire, elle se rencontre essentiellement dans des forêts décidues mais en région méditerranéenne elle fréquente également des habitats sempervirents (chênes verts), des maquis en Corse (voir Blondel 1985, 1986 pour une discussion sur cette particularité) et même des bois de genévriers en Afrique du Nord et des pinèdes aux îles Canaries. La diversité des habitats fréquentés, l’amplitude de l’aire de répartition et l’existence de populations insulaires et de barrières interrompant les flux géniques font donc de cette espèce un modèle de choix pour étudier la variation géographique et d’éventuelles divergences morphologiques et écologiques de certaines populations. Ces divergences peuvent être mises en relation avec certains paramètres environnementaux pour lesquels elles auraient valeur adaptative. Signalons enfin que si cette espèce, ainsi que la Mésange charbonnière (Parus major), ont été tant étudiées, c’est à cause de leur nidification en nichoirs artificiels qu’elles préfèrent même aux cavités naturelles des arbres pour établir leur nid. Ceci facilite la connaissance de paramètres comme la date de ponte, la grandeur de ponte, le nombre de poussins nés et le nombre de jeunes envolés.
L’étude de la variation géographique a donc été pour Darwin un élément important de réflexion dans la mise au point de sa théorie. Nous allons tout d’abord montrer comment de telles études de populations permettent de proposer des modèles de spéciation acceptables. Pour cela, nous allons exploiter un ensemble de travaux menés au Centre L. Emberger (C.N.R.S., Montpellier) par l’équipe de J. Blondel sur la Mésange bleue {Parus caeruleus). L’aire de répartition de cette espèce comprend les îles Canaries, l’Afrique du Nord, la Cyrénaïque, toute l’Europe jusqu’au sud de la Scandinavie au nord et jusqu’à l’Oural à l’est, la Turquie et une partie de l’Iran. Sur l’ensemble de son aire, elle se rencontre essentiellement dans des forêts décidues mais en région méditerranéenne elle fréquente également des habitats sempervirents (chênes verts), des maquis en Corse (voir Blondel 1985, 1986 pour une discussion sur cette particularité) et même des bois de genévriers en Afrique du Nord et des pinèdes aux îles Canaries. La diversité des habitats fréquentés, l’amplitude de l’aire de répartition et l’existence de populations insulaires et de barrières interrompant les flux géniques font donc de cette espèce un modèle de choix pour étudier la variation géographique et d’éventuelles divergences morphologiques et écologiques de certaines populations. Ces divergences peuvent être mises en relation avec certains paramètres environnementaux pour lesquels elles auraient valeur adaptative. Signalons enfin que si cette espèce, ainsi que la Mésange charbonnière (Parus major), ont été tant étudiées, c’est à cause de leur nidification en nichoirs artificiels qu’elles préfèrent même aux cavités naturelles des arbres pour établir leur nid. Ceci facilite la connaissance de paramètres comme la date de ponte, la grandeur de ponte, le nombre de poussins nés et le nombre de jeunes envolés.
APPROCHE BIOMETRIQUE
Une partie du travail de cette équipe, menée par J.L. Martin de 1983 à 1988 (Martin 1988), a consisté à mesurer sur des spécimens de musée (2184 en tout) 18 paramètres morphologiques. Les oiseaux ainsi mesurés provenaient de l’ensemble de l’aire derépartition de la Mésange bleue. Les mesures ont toutes été effectuées par l’auteur. L’analyse d’un jeu de données aussi complexe rend obligatoire le recours à l’outil informatique et à des analyses discriminantes comme l’analyse en composante principale normée (Bouroche & Saporta 1980). Pour simplifier, on peut dire que cette analyse considère l’ensemble des individus étudiés, caractérisés chacun par leurs 18 paramètres, comme des points d’un espace à 18 dimensions. La recherche d’une discrimination entre des groupes éventuels d’individus se fera en projetant ces points dans le plan où le nuage de points sera le plus dispersé. Le programme utilisé indique la contribution de chaque variable à chacun des axes du plan. Cette analyse permet de distinguer un certain nombre de groupes. Elle sépare très nettement les populations eurasiatiques des populations nord-africaines et des Canaries . Dans le groupe eurasiatique, leurs coordonnées sur l’axe vertical séparent des populations continentales et nordiques d’une part (groupes 1, 2 et 3), caractérisées par une taille plutôt grande, des tarses courts et un bec court et trapu, de populations méridionales, péninsulaires et insulaires d’autre part (groupes 4 et 5) caractérisées par une petite taille et un bec légèrement plus fin. Deux oppositions, qui affectent les caractères morphologiques des oiseaux dans le même sens, se superposent ainsi, une qui correspond à un degré d’isolement croissant et l’autre à un gradient latitudinal. On peut remarquer que les populations les plus isolées (groupe 5) sont celles qui se démarquent le plus des autres. Le groupe nord-africain quant à lui se caractérise par des tailles plutôt petites, des tarses longs et des becs longs et fins. On peut remarquer que les différentes populations distinguées par l’analyse au sein du groupe nord-africain sont bien plus différentes entre elles que ce que celles du groupe eurasiatique ne le sont entre elles. Elles sont de plus ordonnées selon un gradient d’isolement de plus en plus marqué.
Peut-on interpréter ces variations en terme d’adaptation ? Tout d’abord l’opposition entre des oiseaux septentrionaux grands et des oiseaux méridionaux petits est en accord avec la règle de Bergmann qui stipule qu’une grande taille, en diminuant le rapport surface/volume, permet de limiter les pertes de chaleur dans des environnements froids chez les espèces homéothermes. On peut également penser que la taille plus forte des individus nordiques est la conséquence d’un pic de ressources alimentaires plus élevé au printemps durant la phase de croissance des jeunes. Concernant la forme du bec, un bec plutôt épais serait mieux adapté à la recherche de nourriture sur les feuilles larges des arbres décidus septentrionaux. Un bec fin se prête par contre davantage à l’exploitation des feuilles plus étroites des essences méditerranéennes ou des aiguilles des conifères dans lesquels niche la Mésange bleue aux Canaries et en Cyrénaïque. Les variations observées dans les caractères de cette espèce semblent donc bien avoir valeur adaptative.
D’un point de vue génétique, cet exemple est très riche d’enseignement car il montre que tant qu’aucune barrière ne s’oppose aux flux géniques, des ruptures marquées ne peuvent pas s’établir entre les populations. Ainsi en Europe, pour les caractères observés, seul un gradient nord-sud est visible. Le passage d’un groupe à l’autre est graduel et l’ensemble des populations reste assez homogène. Les caractères observés expriment dans ce cas la résultante entre les pressions de sélection locales qui tendent à faire diverger ces populations l’une par rapport à l’autre et l’effet unificateur des flux de gènes venus des populations périphériques. Le passage graduel d’un groupe à l’autre exprime le changement en général graduel des caractères de l’environnement. Par contre, dès qu’une barrière géographique s’établit (montagnes ou mer), les populations situées de part et d’autre de cette barrière divergent très nettement l’une de l’autre car l’effet unificateur des flux géniques faiblit ou disparaît. Tout se passe donc comme si cette espèce avait colonisé son aire actuelle de répartition à partir d’un foyer de spéciation dont la localisation reste à préciser. À partir de là, des sous-populations ont pu coloniser d’autres régions dans lesquelles elles ont pu génétiquement diverger, s’adaptant ainsi aux conditions écologiques locales, surtout si des barrières s’opposent à l’arrivée d’autres migrants. D’autres exemples de ce type sont cités par Mayr (1974) qui rapporte que chaque île ou groupe d’îles situé à l’avant d’un continent abrite des variants des espèces continentales.
Dans quel cas ces variants peuvent-ils passer au statut d’espèce à part entière ? Dans sa définition biologique , une espèce est un pool de gènes, protégé des autres pools de gènes, coadaptés par la sélection naturelle pour l’utilisation de certaines ressources du milieu. Une espèce se définit donc par rapport à sa niche écologique (voir plus loin). Les isolats périphériques ne pourront donc être considérés comme de nouvelles espèces que dans la mesure où ils divergent au point de changer de niche. La meilleure preuve de ce changement de statut taxinomique sera la colonisation par des individus de cette nouvelle espèce de l’aire de répartition de l’espèce dont ils sont issus.